



Introduction
Although bamboo is called Daenamu in Korean, where namu means tree, it is a kind of grass belonging to the sub-family of Bambusoideae in the family Poaceae (grasses) that includes 1,670 species in 125 genera globally (Benjamin et al., 2021; Huang et al., 2023). It has been reported that 13 species of bamboo grow in Korea, among these Wangdae (Phyllostachys bambusoides Siebold & Zucc.), Sinidae (Sasa coreana Nakai), and Joritdae (S. borealis [Hack.] Makino) are most common in Korea (Choi et al., 2017).
As in other Asian countries, including China, India, and Japan, which support approximately 80% of the world's bamboo forests, bamboo has been used in Korea for food, building materials, crafts, and high-quality paper as well as for landscaping and soil conservation. For example, fresh and fermented bamboo shoots, known to be high in fat, carbohydrates, protein, minerals, vitamins, enzymes, coenzymes, reducing and non-reducing sugars, and lactic acid, are consumed as foods. Additionally, bamboo salts (jookyeom), regular salt roasted in bamboo culms sealed with yellow clay, and bamboo culms have been used as folk medicine to treat chronic diseases (Lee et al., 2016).
Although bamboo plants normally require little maintenance and can live for decades without much care, insect pests and plant diseases significantly impair the growth and quality of bamboo. Dey et al. (2023) reported that a total of 1,200 insect species, 580 fungi, five bacteria, three viruses, one phytoplasma (mycoplasma-like organism), and one bacterium-like organism have been identified as pathogenic to bamboos. Among these, at least 29 rust fungi belonging to six genera (Kweilingia, Puccinia, Uredo, Phakospora, Stereostratum, and Tunicopsora) cause bamboo rust diseases globally (Nelson and Goo, 2011).
In Korea, flowering and witch's broom disease in bamboo are attracting recent attention, yet information on the bamboo plant is limited. Only two diseases, rust caused by Puccinia longicornis and smut caused by Ustilago shiraiana, are listed in the recent edition of the ‘List of Plant Diseases in Korea’ (The Korean Society of Plant Pathology, 2022). In addition, Yu et al. (2020) reported bamboo as an alternative host of Puccinia spp., which cause rust disease on Chionanthus retusus Lindley, but the pathogenic effect on bamboo species was not discussed.
In April 2022, a yellow fungal mass was observed on the sheath of bamboo, which resulted in the death of bamboo in the flowing year. In this manuscript, we are reporting the causal organism of this disease, which is the first time in Korea to our knowledge.
Materials and Methods
Sample collection and morphological observation.
The fungal sample was corrected from the sheath of the bamboo culm in April 2022. The morphological features of the collected fungal sample were observed by optical microscope (BA410; Motic, Kowloon, Hong Kong) with a Moticam Pro camera. The length and width of spores were measured from 20 randomly selected spores, using the Motic Images Plus 3.0 software.
Genomic DNA extraction, polymerase chain reaction (PCR) amplification, and sequencing.
For genetic identification, genomic DNA was extracted from the teliospores using a 5 min Plant DNA Extraction Kit (Scinomics, Daejeon, Korea). Briefly, a section of the teliospores (<100 mg) was placed in a 2 ml screw-cap tube and 500 μl of DNA solution was added. The spore structure was homogenized using a Bioprep-24 homogenizer (Allsheng, Hangzhou, China) with 2 mm steel beads and 1.00-1.25 mm glass beads, and rotation at 6 m/sec for 30 sec three times. The following process was performed using 400 μl of disruption solution, and the subsequent process was performed, according to the manufacturer's instructions.
The large-subunit ribosomal RNA (rRNA) gene (LSU) sequence, which contains the previously reported sequence region for rust, was amplified using the universal primers, LR0R (5’-ACC CGC TGA ACT TAA GC-3’) and LR7 (5’-TAC TAC CAC CAA GAT CT-3’) for the region containing the previously reported LSU sequence, flanking the conserved sequence region (Aime, 2006). PCR was performed under the following conditions: 95°C for 5 min; 35 cycles of 95°C for 15 sec, 50°C for 15 sec, and 72°C for 1 min 30 sec; and a final cycle at 72°C for 7 min. PCR product was separated on a 1% agarose gel and purified using Expin Combo GP (GeneAll, Seoul, Korea). The purified PCR products were sequenced using Sanger sequencing (Bionics, Seoul, Korea) with LR0R and LR7 primers. The length of the amplified sequence was 1,355 bp, and the sequence was deposited in the National Center for Biotechnology Information (NCBI) database (GenBank accession number OR426436).
Phylogenetic analysis.
The LSU sequence was blasted against sequences in the GenBank database of the NCBI (https://www.ncbi.nlm.nih.gov) using the Nucleotide BLAST program. After sorting by high sequence identity, reference sequences excluding duplicate data were downloaded from the database. The downloaded sequences were aligned using the ClustalW algorithm and trimmed to 1,065 bp in MEGA-X (Kumar et al., 2018). Phylogenetic analysis was performed using the Kimura 2-parameter model and the Bootstrap method with 1,000 replications to produce a maximum likelihood phylogenetic tree. The LSU sequences of three species, Gymnosporangium juniperi-virginianae AFTOLID 712 (AY629316.1), Kuehneola uredines AFTOLID 987 (AY745696.1), and Insolibasidium deformans AFTOLID 722 (AY646099.1), were downloaded from NCBI database and were used as outgroup in phylogenetic analysis.
Results
Morphological observation.
The color of the fungal sample collected was yellow, and it was observed that the sheath of the bamboo culm was damaged (Fig. 1A-C). The sample collected was considered to be at the sporophytic stage in the life cycle of Pucciniales (Aime et al., 2018). After about 3 months, the infected bamboo no longer had yellow telia, and the upper side of the bamboo tree from the fungal infection of the bamboo had died (Fig. 1C). The leaves of uninfected bamboo were fresh green; however, the leaves of infected bamboo turned brown and died in the following year (Fig. 1D). In this note, we report the causal organism of the observed disease, which to the best of our knowledge is the first report of the disease in Korea.
Fig. 1.
Symptoms of bamboo tree infected with Stereostratum corticioides. (A) Pseudosasa japonica culms with telia of S. corticioides. (B) Picture of telia developed on the culms was taken on April 30, 2022. (C) Picture of the same infected area was taken on 29 July, 2022. Discoloration of the culms and sheath of the infected site is different from underneath that was not infected. (D) Population of P. japonica showing top blight symptoms 1 year after observation of telia on culms.

Microscopic observation of the collected samples showed that the yellow structures on the outside of the bamboo culm were teliospores with a pedicel attached to one end (Fig. 2). The spores were observed as oval in shape, brown in color, and possessing a one-septate structure (i.e., two-celled; Fig. 2). The length and width ranges of 20 random spores were 25.70-39.6 μm and 18.23-23.94 μm, respectively, and the average length and width were 31.83±3.57 μm and 20.74±1.72 μm, respectively. These measurements differed by only about 5% from the first reported length of 33.6 μm and width of 22.0 μm for 10 spores of Stereostratum corticioides (Okane et al., 2020). While teliospores predominate, some urediniospores were identified, which formed in the uredinium, a stage before the formation of the telium.
Fig. 2.
Morphological characteristics of Stereostratum corticioides spores. The oval-shaped telia, which accounts for the most of, with one-septate separated from the spore crowd, was thought to be a yellow mass. The red arrows indicate the urediniospores. Microscopic images were observed without any staining at 200× (left) and 400× (right) magnification, respectively.

Identification of the rust fungi.
In order to identify the yellow mass on bamboo culm, the LSU rRNA gene was amplified and the nucleotide sequence was analyzed. The length of the analyzed sequence after amplification was 1,355 bp and the sequence was deposited in the NCBI database (OR426436).
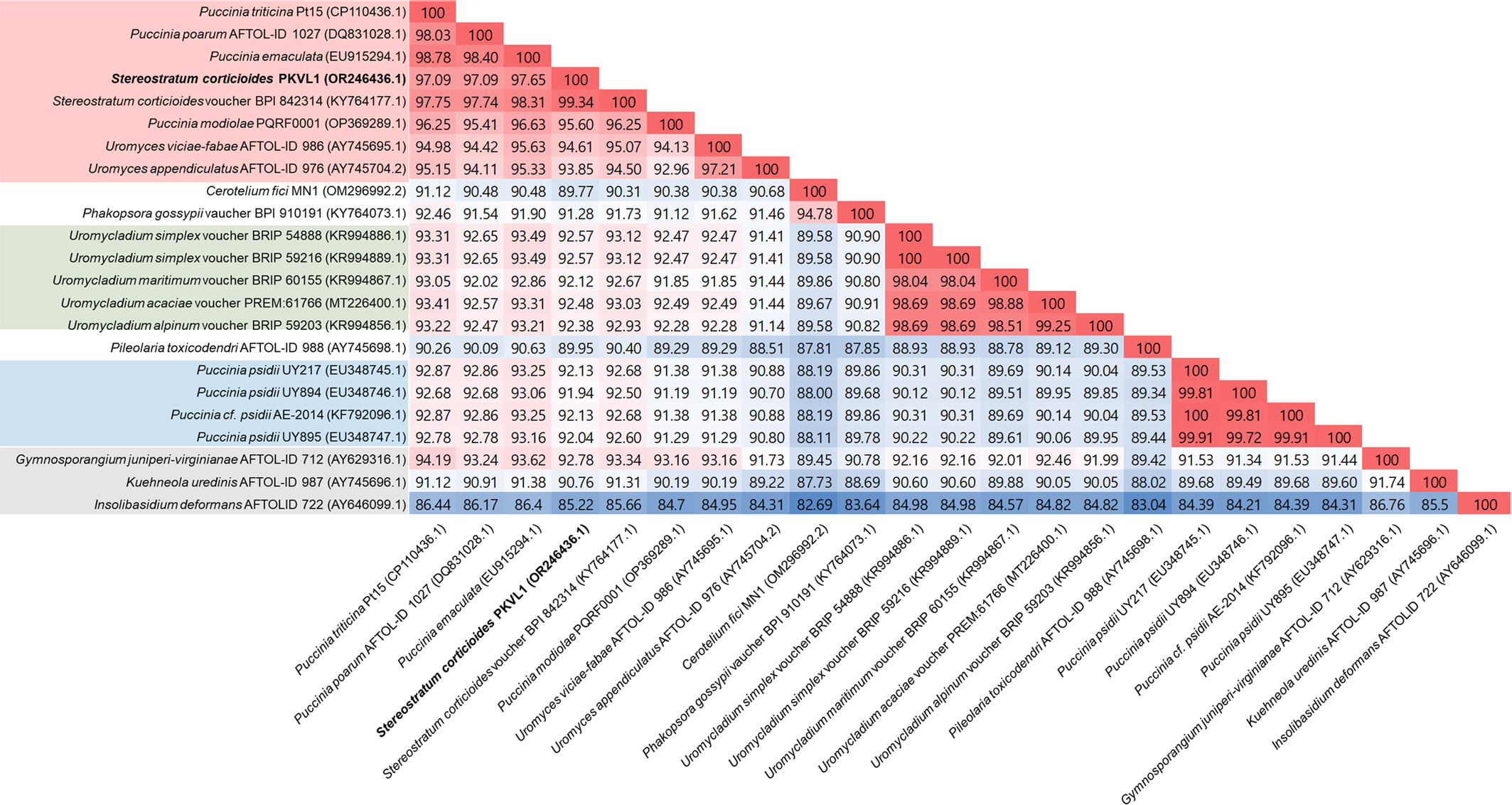
The nucleotide sequence showed the highest sequence identity of 99.36% to Stereostratum corticioides voucher BPI 842314 (Fig. 3; GenBank accession number KY764177) and high sequence homology to P. emaculata (97.79%), P. poatum AFTOLID 1027 (97.42%), and P. triticina Pt15 (97.28%; Fig. 3). Phylogenetic tree analysis showed that the strains clustered into three groups of high similarity (a closed group with S. corticioides; the genus Uromycladium; and P. psidii strains), while some strains did not cluster into any of those groups, such as Cerotelium fici MN1, Phakopsora gossypii vaucher BPI 910191, Pileolaria toxicodendri AFTOL-IF 988, Gymnosporangium junipiri-virginianae AFTOL-IF 712, Kuehneola uredines AFTOLID 987, and Insolibasidium deformans AFTOLID 722 with homologies of 89.77%, 91.28%, 89.95%, 92.78%, 90.76%, and 85.22%, respectively, based on trimmed sequences (Figs. 3, 4).
The five strains belonging to Genus Uromycladium form their own clade and were found to be closely related to Cerotelium fici MN1 and Phakopsora gossypii vaucher BPI 910191, which belong to Phakopsoraceae family, in the phylogenetic tree, but their LSU sequence similarity was only about 89% (Figs. 3, 4). Interestingly, four P. psidii strains were placed in a separate clade to the closed group with S. corticioides in which three other Puccinia species (P. triticina Pt15, P. poarum AFTOLID 1027, and P. emaculata) were placed (Fig. 4). The sequence similarity of P. psidii strains to the other three Puccinia species ranged from 92.68% to 93.25%, which was lower than that of S. corticioides strains (97.09-98.31%) and Uromyces species (94.42-96.63%; Fig. 4). These differences in homology lead to different positions of the branches between S. corticioides and Puccinia species in the phylogenetic tree (Fig. 3).
Fig. 3.
Phylogenetic tree of Stereostratum corticioides by maximum likelihood using LSU gene sequence. The numbers above the nodes are the supporting percentages obtained from 1,000 bootstrap replicates. Only bootstrap values above 80 are shown. The National Center for Biotechnology Information GenBank accession numbers are indicated in parentheses. The scale bar represents a 2% nucleotide sequence divergence. Red background, a group related to Puccinia species that does not include P. psidii; Green background, Uromycladium species group; Blue background, P. psidii strains group; Gray background, outgroup.

The homology of LSU from S. corticioides PKVL1 to Uromyces viciae-fabae AFTOLID 986 and U. appendiculatus AFTOLID 976 was 95.60% and 94.61%, respectively, which was higher than the homology to P. modiolae (93.85%; Fig. 4). The fact that Puccinia species, S. corticioides, and Uromyces species cluster closely together in the phylogenetic tree indicates high genetic similarity (Figs. 3, 4). The genetic relationship was complicated because of the similarity of the homology between the Puccinia species and Uromyces species, but S. corticioides was clearly branched into one of them (Fig. 3).
Discussion
Despite the economic and ecological importance of bamboo species, diseases in bamboo have been little studied in Korea, and to date only two fungal diseases have been officially recorded. A mass of yellow fungal growth was observed on the culm of a bamboo plant in the spring of 2022, and microscopic observation indicated that it was the telia of a species of Puccinia. Subsequent molecular characterization of the LSU rRNA gene indicated that the fungus has a 99.36% sequence identity to S. corticioides. The only member of the genus Stereostratum in the family Pucciniaceae is S. corticioides, which was reassigned from Puccinia corticioides to S. corticioides based on the morphological features including coriaceous telia with many bicellular teliospores (Magnus, 1899). However, nucleotide sequence analysis of the internal transcribed spacer 2 gene of rDNA and the D1-D2 region of the large-subunit rRNA gene (LSU) of S. corticioides and 10 strains of P. corticioides resulted in 99% bootstrapped values using neighbor joining and maximum likelihood methods and a Bayesian inference posterior probability of 0.80 (Okane et al., 2020). Consequently, Okane et al. (2020) proposed reviving the binomial P. corticioides to replace S. corticioides.
Fig. 4.
Similarity heatmap of LSU sequences of Stereostratum corticioides and related species. Red indicates relatively high homology (100%), and blue indicates relatively low homology (82%).
P. corticioides has been reported from China, Japan, India, Hawaii, Pakistan, and Taiwan (Cummins and Kimura, 1971; Gardner and Hodges Jr, 1989; Hiratsuka, 1992; Katumoto, 1968; Spaulding, 1961; Tangjang et al., 2018). In Korea, only one rust fungus, Puccinia longicornis has been reported as the causal organism of rust on bamboo species including Phyllostachys spp., Pseudosasa japonica, and Sasa quelpaertensis, and this is the first report of culm rust caused by S. corticioides (Lee, 2001). The teliospores of S. corticioides have an oval shape with no extrusion at the end (Fig. 2) unlike the teliospores of S. longicornis, which are fusiform or cylindrical-fusiform with a pointed extrusion. Furthermore, molecular analysis of the LSU genes confirmed that the causal organism is S. corticioides.
The spermogonia and aecia of P. longicornis in Korea have not been characterized. However, nucleotide sequence analysis of a rust fungus on Choerospondias axillaris (Anacardiaceae) in Japan confirmed the fungus was the aecial stage of P. corticioides (Okane et al., 2020). Although C. axillaris does not occur naturally in Korea, five species of the Rhus genus of the Anacardiaceae family including R. javanica L. do occur in Republic of Korea. Therefore, further research on the etiology of S. corticioides, including host range and the possible impact on bamboo forest in Korea, is required.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






