


Introduction
Permafrost soils are reservoirs of diverse uncharacterized microbes, including plant pathogens (Wu et al., 2022). The emergence and spread of novel plant pathogens may threat-en the global food supply. The subarctic permafrost has experienced a temperature increase of 2-3°C since the 1970s (Scheel et al., 2022). Recent rapid warming has extended the permafrost thawing period and soil depth of active layer, resulting in a high incidence of new pathogen emergence (Ng et al., 2014; Yarzábal et al., 2021). To date, plant pathogen outbreaks in melted permafrost have not been reported and remain largely unexplored. Therefore, our knowledge of potentially harmful microbes in permafrost soils during summer remains insufficient. Rising temperatures, influenced by global climate change, are expected to activate these microbes, which were previously dormant in the permafrost. Understanding its emergence and pathogenicity in permafrost soils is critical for maintaining a stable food supply.
Bacterial soft rot pathogens are a major cause of losses in vegetable production and significantly impact both food security and sustainability. They are harmful at all stages of fruit and vegetable production because they secrete plant cell wall-degrading enzymes, such as pectinases, cellulases, and proteases, and cause tissue maceration (Toth et al., 2003). In the field production of potatoes, soft rot pathogens cause economic losses during harvesting, storage, and transportation processes. Bacterial soft rot pathogens are diverse and ubiquitous and can spread asymptomatically in the environment. The genera Dickeya and Pectobacterium, belonging to the family Pectobacteriaceae, are the most extensively studied bacterial soft rot pathogens and have caused constant problems in vegetable production, particularly in the case of potatoes (Charkowski, 2018; Park et al., 2023). In addition to the Dickeya and Pectobacterium species, mesophilic and psychrotolerant (cold-adapted) Pseudomonas strains of the family Pseudomonadaceae have recently emerged as significant pathogens affecting carrots, onions, lettuce, melons, spinach, and potatoes (Charkowski, 2018; Godfrey and Marshall, 2002; González et al., 2003; Kim et al., 2002; Wasendorf et al., 2022; Zhang et al., 2016). A complex infection by various bacterial pathogens from different families, such as Pectobacteriaceae and Pseudomonadaceae, can exacerbate vegetable soft rot disease. Unfortunately, very little information is available on Pseudomonas pathogens emerging from cold environments including subarctic regions, and the virulence mechanisms of soft rot have not yet been studied.
Materials and Methods
Description and culture conditions of bacterial strains.
The psychrotolerant bacterium N3-W was isolated from tundra soil in Alaska, USA in 2019. Based on the whole genome and 16S rRNA sequence identities, this isolate could not be categorized at the species level in the genus Pseudomonas; thus, it was named Pseudomonas sp. N3-W (=KACC 22935) (Kim et al., 2022). Pectobacterium carotovorum KACC 16999 and Pseudomonas syringae pv. tomato KACC 15103 were used as the pathogenicity-positive strains for soft rot of potato tubers and hypersensitive necrotic spots on hot pepper leaves. For physiological and pathological tests, the bacterial strains in 20% glycerol stock were transferred onto R2A (KisanBio, Seoul, Korea) agar plates, incubated at 25°C for 2-3 days, and stored at 4°C until used.
Growth measurement at different temperatures.
Strains N3-W and KACC 16999 were cultured in liquid R2A medium at 15°C for 2 days with shaking and then transferred to new 50 ml R2A adjusting to an optical density at a wavelength of 600 nm (OD600) of 0.01. The cell suspensions were cultured at 5°C, 15°C, or 25°C for 4 days. During culture at each temperature, the OD600 values were spectrometrically measured at the time interval of 1 day. At the same time, the bacterial growth was tested on R2A plates by incubation at 5°C, 15°C, or 25°C for 4 days.
Enzyme activity determination.
The glycerol stock of strain N3-W, KACC 16999, or KACC 15103 was transferred onto R2A agar plates and incubated at 25°C. The grown colonies were picked and then streaked using a disposable plastic inoculation loop on R2A plates containing 1.0% (w/v) skim milk (BD, Difco, Sparks, MD, USA) and incubated at 25°C for 3 days. Proteolytic activity was arbitrarily scored according to the size of the clear zone surrounding the colonies.
The bacterial strains were incubated on R2A plates containing 1.0% (w/v) carboxymethyl cellulose (C4888, Sigma-Aldrich, St. Louis, MO, USA) at 25°C for 7 days. The cells were stained with 5 ml of 0.1% (w/v) Congo red solution for 20 min and destained twice with 1.0 M NaCl for 15 min. The cellulolytic activity was scored according to the size of the clear zone (light orange).
The bacterial strains were incubated on R2A plates containing 0.1% (w/v) citrus peel pectin (P9135, Sigma-Aldrich) at 25°C for 3 days. The grown cells were stained with 0.5 mM iodine solution and incubated for 20 min. The plates were slowly washed with water. The pectinolytic activity was scored according to the size of the clear zone (yellow).
Hypersensitivity test on hot pepper.
To test for hypersensitivity reactions by potential plant pathogens, strains N3-W and KACC 15103, grown in liquid R2A at 15°C, were washed with water to remove the remaining medium and diluted in Murashige and Skoog (MS) medium to OD600 values of 0.6. The cell suspension was inoculated into the leaves of hot pepper seedlings (Capsicum sp. cultivar Maepgona) using the syringe pressure infiltration method. Negative control plants were inoculated with the same volume of 1/2-diluted MS medium (1/2 MS). The inoculated peppers were placed in a dew chamber at 23°C (16 hr of light and 8 hr of darkness) and 70% relative humidity for 3 days. After the appearance of necrolytic spots in the inoculated leaves, bacterial colony-forming units (cfu) were measured on R2A plates incubated at 25°C using a conventional serial dilution method.
Pathogenicity test on potato tuber at different temperature regimes.
Potato tubers (Solanum tuberosum) were surface-sterilized by dipping in 1.0% sodium hypochlorite solution for 15 min, washed twice with sterile distilled water, and left on a table to air dry overnight. After the potato surfaces were dried, slices of approximately 2.0 cm width were cut using a sterilized knife and transferred to petri dishes containing 70-mm-diameter filter paper dampened with water. The bacterial inocula of N3-W and KACC 16999 were prepared following the same procedure used for the hot pepper leaves. The cell suspension (50 μl; final OD600 value of 0.6) was inoculated onto the slices. The control was inoculated with the same volume of 1/2 MS. The inoculated potatoes were placed in an incubator at 5°C, 15°C, or 25°C for 3 days.
Results and Discussion
Growth measurement at different temperatures.
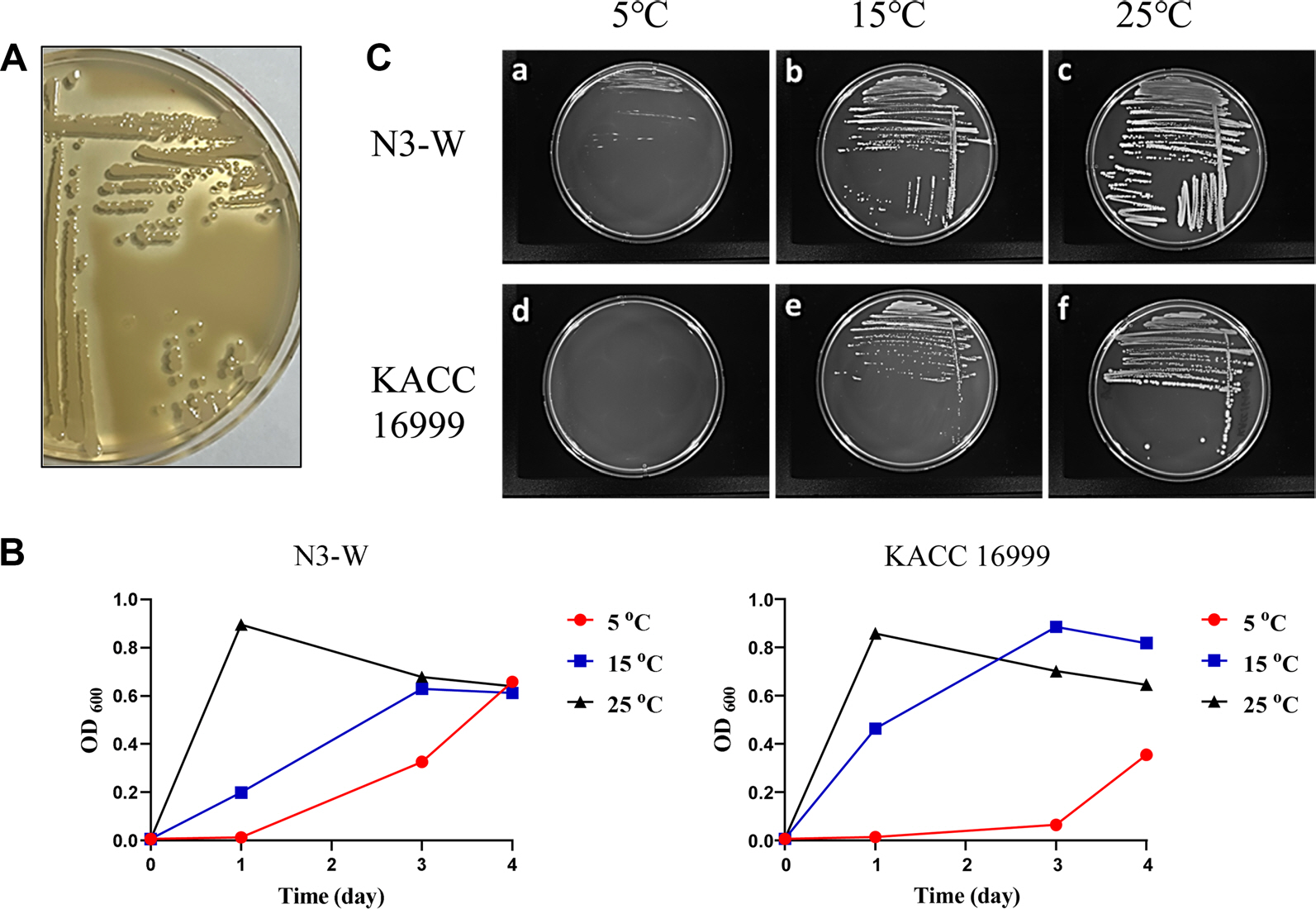
Psychrotolerant N3-W was characterized to show strong proteolytic activity on skim milk-containing agar plate at a temperature range of 5-25°C (Fig. 1A). The comparison of cell growths of N3-W and mesophilic KACC 16999 in liquid R2A was performed by measuring the OD600 values during culturing at different temperatures (5°C, 15°C, or 25°C) (Fig. 1B). The growth of N3-W cells significantly increased when cultured at 25°C, with maximal growth after 1 day of culturing. Maximal growth at 15°C was reached after 3 days, while growth at 5°C increased slowly and continuously until day 4. The cell growth patterns of KACC 16999 were similar to those of N3-W, starting to increase shortly after incubation at 25°C and reaching the maximal growth after 3 days of culturing at 15°C. In contrast, its growth was significantly delayed compared to that of its psychrotolerant counterpart, N3-W. When the colony growth of strains N3-W and KACC 16999 was examined on R2A plates, the growth rates were similar to those of liquid cultures at all tested temperatures (Fig. 1C). These results indicate that N3-W is metabolically active and can grow at the low temperatures of refrigerating compartments.
Fig. 1.
Growth characterization of N3-W. (A) Clear zone production around N3-W colonies owing to its proteolytic activity on skim milk plate. (B, C) comparisons between the growth rates of psychrotolerant N3-W and mesophilic KACC 16999 at different temperatures of 5°C, 15°C, and 25°C. The growth of N3-W was more distinct than that of KACC 16999 at 5°C.
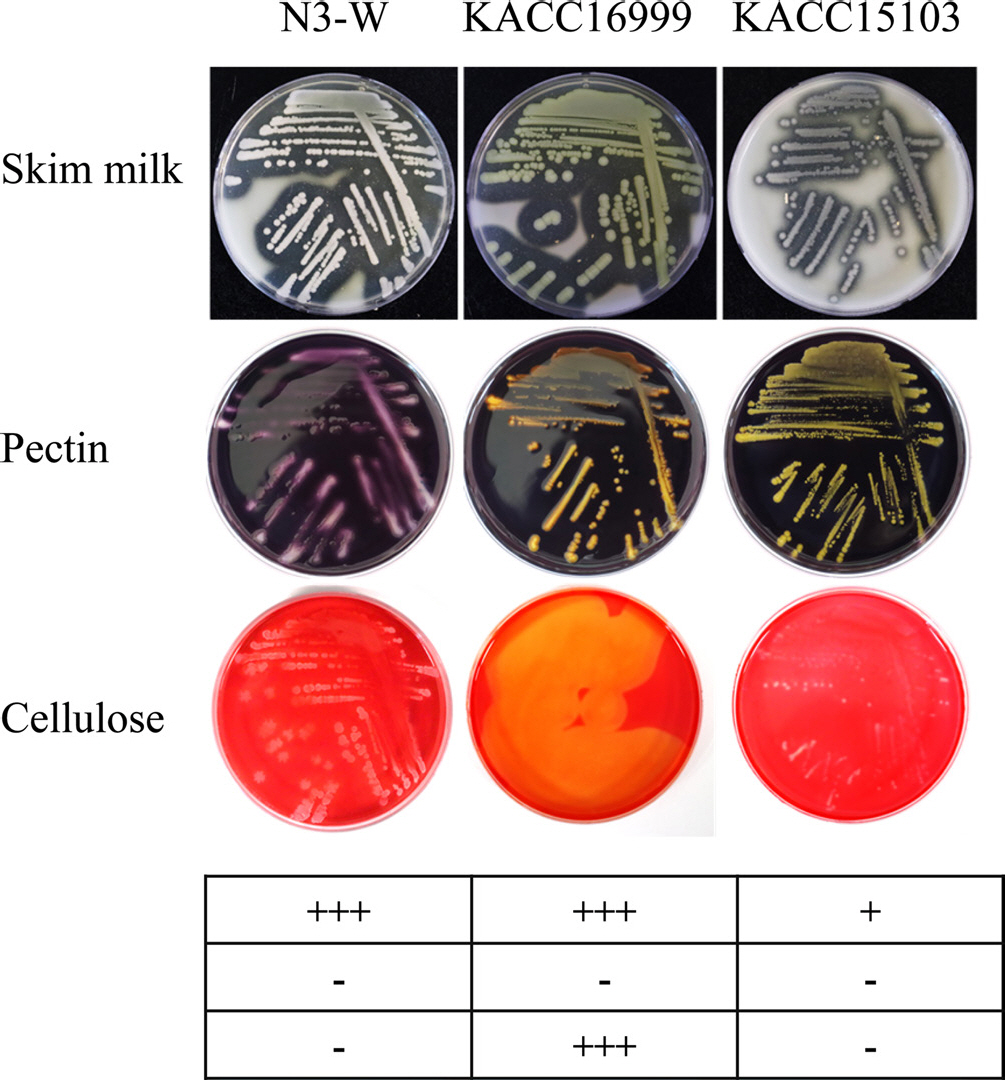
Hydrolytic enzyme activity for plant cell wall polymers.
Generally, bacterial soft rot is caused by plant cell wall decomposition by various enzymes, such as proteases, cellulases, and pectinases. Therefore, we tested the ability of N3-W, KACC 16999, and KACC 15103 to produce hydrolytic enzymes using the plate method. Enzyme activity was tested using skim milk, carboxymethyl cellulose, and pectin as surrogate substrates to evaluate the hydrolytic activities of proteases, cellulases, and pectinases, respectively. N3-W and KACC 15103 produced proteases, but not cellulase or pectinase, while KACC 16999 showed strong protease and cellulase activities, but not pectinase activity (Fig. 2).
Fig. 2.
Enzymatic activity tests on agar plates containing each plant cell wall polymer at 25°C. Clear zone production around bacterial colonies owing to the proteolytic activity for skim milk, pectolytic activity for pectin, or cellulolytic activity for cellulose was compared among psychrotolerant N3-W and mesophilic KACC 16999 and KACC 15103. The enzymatic activity is indicated with symbols below the photo: +++, strong; ++, medium; +, weak; -, inactive.
Hypersensitivity induction on hot pepper leaf by N3-W.
The N3-W cell suspension was infiltrated into the leaves of hot peppers to test for hypersensitivity reactions induced by the virulence of potential plant pathogens. The positive control KACC 15103 and the suspected pathogen N3-W produced brown necrotic spots on the inoculated pepper leaves after 3 days of inoculation, with N3-W inducing larger necrotic regions than KACC 15103. In contrast, the mock control (without inoculation) did not induce symptoms (Fig. 3A). The cfu values of KACC 15103 and N3-W from each inoculated pepper leaf were approximately 18(±5)×105 cfu/ml and 314(±14)×105 cfu/ml, respectively, whereas no bacterial colonies were found with the mock control (Fig. 3B). These experiments indicated that N3-W is a plant pathogenic bacterium that can infect and proliferate in pepper leaves.
Fig. 3.
(A) Bacterial necrotic spots induced on red pepper leaves after inoculation with N3-W and KACC 15103 (positive control). Mock denotes healthy leaves treated with 1/2 MS (negative control). Bioluminescence images of necrotic spots were taken with the Azure Imaging System (Azure Biosystems, USA). (B) Bacterial colonies from the necrolytic spots induced after inoculation with N3-W or KACC 15103.

Pathogenicity test on potato at different temperature regimes.
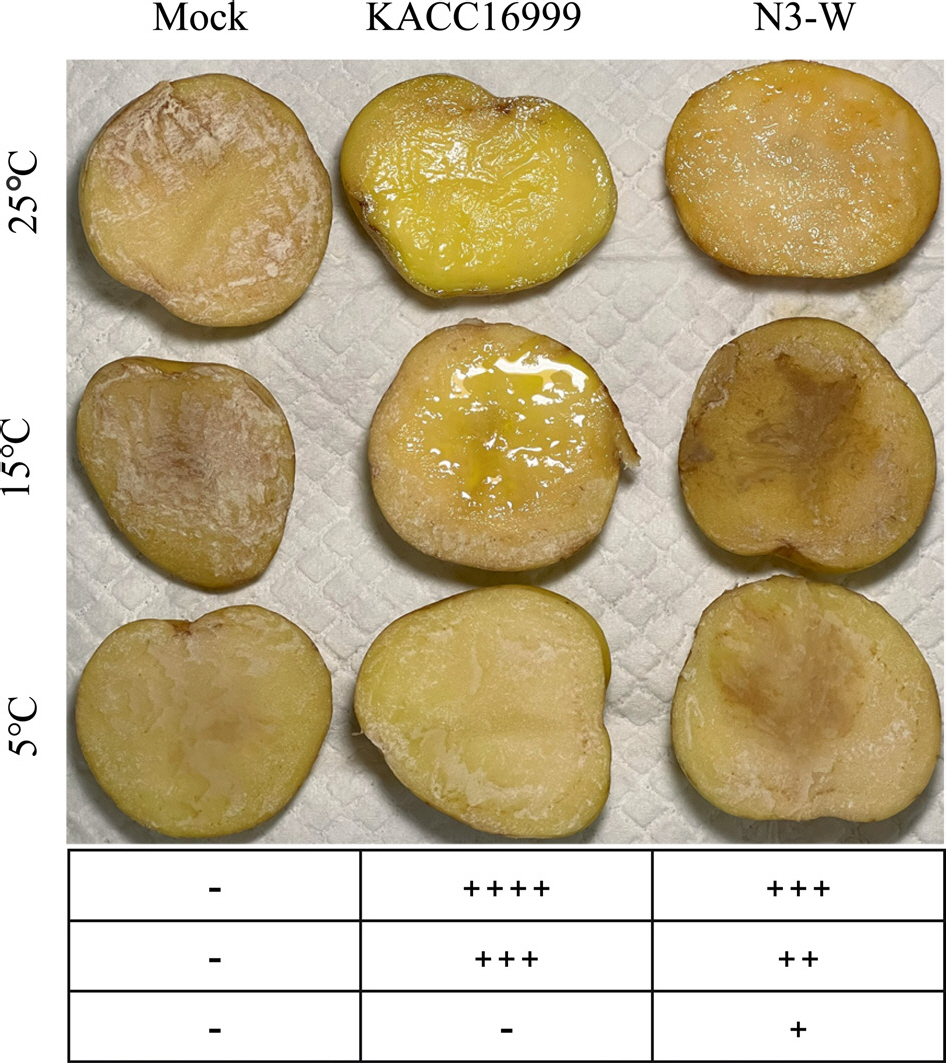
The cell suspension of N3-W was inoculated onto potato slices and incubated at different temperatures for 3 days, to evaluate potato soft rot disease. In accordance with previous test results that N3-W has biological activity at a broad temperature range of 5-25°C, with an optimal temperature of 25°C, it caused bacterial soft rot symptoms at 5 (weak), 15 (medium), and 25°C (strong). The positive control KACC 16999 caused a more severe disease at 15°C (strong) and 25°C (very strong) than N3-W. In contrast to N3-W at 5°C, KACC 16999 did not induce soft rot symptoms, matching its lower biological activity than N3-W at the low temperature of 5°C (Fig. 4).
Fig. 4.
Bacterial soft rot symptoms induced on potato tubers after inoculation with N3-W and KACC 16999 (positive control). Mock denotes healthy tubers treated with 1/2 MS (negative control). The soft rot symptoms were photographed after a 3-day incubation of inoculated potatoes at 5°C, 15°C, or 25°C. The severity of soft rot on potatoes is indicated with symbols below the photo: ++++, very strong; +++, strong; ++, medium; +, weak; -, nonpathogenic.
Generally, bacterial soft rot is caused by cell wall decomposition by various enzymes such as pectinase, cellulase, and protease. N3-W showed strong proteolytic activity, but no pectinolytic or cellulolytic activity, and produced soft rot symptoms in potato tubers. Using the N3-W whole-genome sequence information, nine genes were assigned to various extracellular proteolytic enzymes, including metalloproteases and serine proteases. Among them, one (GenBank ID: UWF51570.1) was identified as a potential virulence factor with high similarity to serralysin family metalloproteases from Pseudomonas spp. Except for the proteases, we could not detect the genes for plant cell wall-degrading enzymes (i.e., pectinases and cellulases), which directly contribute to the maceration of plant cells and tissues. Remarkably, the other subarctic Pseudomonas isolates causing soft rot in potatoes produced only protease (unpublished data), indicating that protease could function as the sole virulence factor for potato soft rot caused by psychrotolerant bacteria.
The number of farms in Alaska has been on the rise, increasing from 440 in 2014 to 590 in 2021, making it the top state in terms of new farm growth in the United States (Mertz and Benz, 2022). This increase in farms is believed to have accelerated significantly since 2019, likely because of the coronavirus disease 2019 pandemic. Rising temperatures are expected to activate soft-rotting soil bacteria previously dormant in frozen soil. Our research highlights the importance of studying the spread and pathogenicity mechanisms of vegetable soft rot pathogens, which will become more active owing to soil warming and melting.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






