



ì ëĄ
êČœì íë „ê°ë°êž°ê”Ź(Organization for Economic Cooperation and Development)ì ë°ë„Žë©Ž ìì ì ìŽêł ì§ìê°ë„í ìë볎êžì ìíìŹ íìŹ ëëč 1.5ë°° ìŽìì ìëŹŒì ìì°ì±ì ìê”Źíêł ìë€(JĂ€germeyr ë±, 2020; Xu ë±, 2019). 2050ë
êčì§ ìžêł ìžê”Źê° ìœ 97ì”ëȘ
ì ìŽë„Žêł êž°íì êŽë šë ìëŹŒ ìì°ì±ìŽ ê°ìí êČìŒëĄ ììëìŽì ž ìì ì ìŽêł ì§ìê°ë„í ëì
ì ìëì볎 돞ì ëĄ ìžìëêł ìë€(Chen ë±, 2016; Lau ë±, 2017; Rodriguez ë±, 2019). ìŽëŹí êŽì ë€ì ìžë„ê° íìëĄ íë ëì
ì ë
čì íëȘ
ì ê°ííêł phytobiome ê°ë
ì ëì
ì쌰ë€. Phytobiomeì 믞ìëŹŒ ìžìì ê°ìŽ ìëŹŒì 볎ížíêł , ìì°ì±ì ëìŽêž° ìí ìŁŒëł ìžìê° ìíž ì ì”í ë°©í„ìŒëĄ ìì©íë©° ìëŹŒìŹë°°ì êŽë šë ëȘšë ìžìë„Œ ì믞íë€(Xiong ë±, 2021).
ê·ž ì€ ìëŹŒ êŽë š 믞ìëŹŒì í ìë¶í° ìëŹŒì íêž°êčì§ ìëŹŒì ëȘšë ìĄ°ì§ì 믞ìëŹŒì ììĄŽ êł”ê°ìŒëĄ ìŹì©íêł ììŒë©° ê°ê°, ê·Œê¶(rhizosphere), ëŽê¶(endosphere), ìœê¶(phyllosphere) ë° ìą
ììëŹŒê”°êł(seedbiome)ëĄ ë¶ë„ëë€(Bintarti ë±, 2022; Hannula ë±, 2019; Kim ë±, 2019a; Lu ë±, 2018). ëí ì§ë ììë
ëì ì°ê”ŹëìŽ ìš plant growth-promoting rhizobacteria (PGPR) êŽë š 믞ìëŹŒì ìëŹŒì ⧠ëčìëŹŒì ì€ížë ì€ëĄ ìží íŒíŽë„Œ ìŒì ë¶ë¶ êČœê°ìíŹ ì ìë êČìŒëĄ ìë €ì ž ìë€(Berkelmann ë±, 2020). PGPR 믞ìëŹŒì êŽë š êž°ììŒëĄ IAA (indole-3-acetic acid) ìì°, ì§ì êł ì , siderophore ìì° ë± ë€ìí êČœëĄë„Œ í”í ìëŹŒ ìì„ ìŽì§ ë„ë „ì ê°ì§êł ììŒë©° ëíì ìž ëŻžìëŹŒ ê”°ìŒëĄ Bacillus, Pseudomonas ê·žëŠŹêł Streptomycesê° ììŒë©° ìŽë€ì ìëŹŒì ìì„ì í„ììí€ë ì§ê°ì ë©ì»€ëìŠì ê°ì§ êČìŒëĄ ìë €ì ž ìë€(Andleeb ë±, 2022; Bokhari ë±, 2019; Fidan êłŒ Zhan, 2019; Herrera-Quiterio ë±, 2020; Oviedo-Pereira ë±, 2022).
ê·ž ì€ Streptomyces ìì ìëŹŒì ê·Œê¶, ìœê¶ ê·žëŠŹêł ëŽê¶ìì ê”°ì§íê° ê°ë„í êČìŽ íìžëìë€(Lindowì Brandl 2003; Redford ë±, 2010). ëí 1940ë
actinomycin ë°êČŹ ìŽí, ìëŹŒíì íì±ì ê°ì§ íìì ë° ì ì© 2ì°š ëìŹì°ëŹŒì í볎ì í”ìŹ ëŻžìëŹŒëĄ ìžì§ëêł ìë€(Bentley ë±, 2002; Deshpande ë±, 1988; Lee ë±, 2020). íìŹêčì§ ìë €ì§ íìì ìŽ 70-80%ì íìì ê° Streptomycesììì ë¶ëŠŹëìŽ ëłŽêł ëìë€(Harir ë±, 2018). Streptomyces ìŽì°šëìŹì°ëŹŒêłŒ êŽë šë êž°ìĄŽì ì°ê”Źì ë°ë„Žë©Ž, Streptomycesì ìíì ëšêł ì€ êž°ê· ìŹ(aerial hypha)ëĄë¶í° íŹìê° íì±ëë ëšêłìì ìŽì ëšêłìì ìì±ëì§ ììë 2ì°š ëìŹì°ëŹŒì ì íìŽ ìŽëŁšìŽì§ëŻëĄ íŹìì íì±ì ë°©ì ê· ì íìëŹŒì§ ìì°ì ììŽ ì€ìí êłŒì ìŒëĄ ìë €ì ž ìë€(YagĂŒe ë±, 2012).
ì íì°ê”Źìì í볎ë ì€ëŹŒ ìì Streptomyces globisporus SP6C4ë ëžêž° ìë€ìëłêłŒ íêž°ìì ë°ìíë ìżëčêł°íĄìŽëłì ëíìŹ ì ì©ëŻžìëŹŒëĄ ì íčì±íëììŒë©°, ìŽë ëì ìëŹŒëł ë°ì ì”ì ì SP6C4ê· ìŁŒê° ìì°íë ëŽìŽì± íí©ëŹŒêłŒ ìì©ìŽ êŽìŹíšìŽ ë°íìĄë€(Cha ë±, 2016; Kim ë±, 2021). S. globisporus SP6C4ë 2ê°ì§ì ì ê· lantipeptideë„Œ íŹìíì± ëšêłìì ìì°íë©°, ìŽë conprimycinêłŒ grisinìŒëĄ ì§ê· ìžíŹëČœ ìí©ì± ë° RNA ì€í©íšì íì±ì ì”ì íìë€(Cha ë±, 2016; Kim ë±, 2019b). ëí Streptomyces sp. S8ì ìë ìŹë°°ì§ ë° ìŹì©ì§ìì ë°êČŹëë large patch(ê°ìë§ëŠëł)ë„Œ ë°ììí€ë ëłìê· Rhizoctonia solani AG2-2ì ìì„ì ì”ì í ì ìë íìí êžží íčì±ì ê°ì§êł ìë êČìŒëĄ ëłŽêł ëìë€(Jeon ë±, 2019). Streptomyces sp. S8ì genome sequencing ì ëłŽë„Œ ë°íìŒëĄ CRISPR/Cas9 systemì í”íŽ valinomycin ìí©ì± ì ì ì ìì ëì°ëłìŽë„Œ ì ìì í”íìŹ, íŽëč ëì°ëłìŽë large patch ëłìê· ì ëí íì§ê· íì±ìŽ ìì€ëë êČì íìžíìë€(Jeon ë±, 2019).
ìëŹŒëłì ë°ìì íšêłŒì ìŒëĄ ì”ì íë êž°ë„í ì ì©ëŻžìëŹŒì ì€ì©íë„Œ ìíìŹ ëë ë°°ì êł”ì ìŽ ë°ëì íìíë€(Park ë±, 2018, 2019). íì§ë§ 믞ìëŹŒì ììë°°ìì ìŁŒëĄ êł ê°ì ì€íì© ë°°ì§ê° ìŹì©ëëŻëĄ ëì
íì„ìì ìŽë„Œ íì©íêž°ìë ìì§ ë§ì íêłì ìŽ ìĄŽìŹíë€(Park ë±, 2018). ëí ì ì©ëŻžìëŹŒ ì ì ëĄ íì±ì ëíëŽë©° ìíëêž°êčì§ë ë§ì êł”ì ìŽ íìíë©°, ê·žì€ ë°©ì ê· ì ë°°ìì ììŽ êčë€ëĄìŽ íčì±ìŽ ììŒëŻëĄ íìŹêčì§ ë°°ì ìĄ°ê±Žì í늜íë êČìŽ ì°ì ëìŽìŒ íë€(Jung ë±, 2011). ëì
믞ìëŹŒì ëëë°°ìì, ìŒë°ì ìŒëĄ ë°°ìëì 믞ìëŹŒì ìì„ìŽ ìŽì§ëë íìì, ì§ììì ëí ì ì ìŽ ê°ì„ ì€ìíë€. ëí ë°°ì§ì ì°ë, ë°°ììšë, ë°°ìêž°ì íì ì ë±ìŽ êł ë € ëììŽë€(Lee ë±, 2021). ëłž ì°ê”Źììë, íì§ê· ëŹŒì§ìŽ ê”ŹëȘ
ë 2ìą
ì ì ì© ë°©ì ê· ì íì©íìŹ ê°ëł 믞ìëŹŒ íčìŽì ìŒëĄ ìì„êłŒ íŹìíì±ì êŽìŹíë ëŹŒì§ì ê”ŹëȘ
íêł ì íìë€. êž° ì ì ë ì ì©ë°©ì ê· 2ìą
ì ëëìì°êł”ì ì íìí íììêłŒ ì§ììì ê·ëȘ
íë êž°ìŽì°ê”Źë„Œ ë°íìŒëĄ ìëŹŒíì ë°©ì ì ê°ë°ì ëí êž°ìŽ ìëŁë„Œ ë§ë šíêł ì íë€.
ìŹëŁ ë° ë°©ëČ
PGPR íšì íì± êČìŠ
Streptomyces sp. S8ë 2017ë
ìë ìŹë°°ì§ì ê·Œê¶ìì ë¶ëŠŹëììŒë©°, ì§íë¶ ì§ê· ëłìê· ì ëí ì°ìí íì§ê· ë „ì ê°ì§êł ìë€(Jeon ë±, 2019). S. globisporus SP6C4 ê· ìŁŒë ëžêž° êœìì ë¶ëŠŹë ë°©ì ê· ìŒëĄ ì§ìë¶ ì§ê· ëłìž ìżëčêł°íĄìŽëł, êœêł°íĄìŽëłì ë°ìì íšêłŒì ìŒëĄ ì”ì íë íčì±ìŽ ëłŽêł ëìë€(Kim ë±, 2019a). Streptomyces sp. S8êłŒ S. globisporus SP6C4ì íŹì ííìĄ ì ìĄ°ë„Œ ìíŽ MS media (mannitol 20 g, soya flour 20 g, agar 20 g per liter)ì ëë§íìŹ 28°C ë°°ìêž°ìì 7ìŒê° ë°°ìíìŹ íŹìë„Œ íì±íìë€. ë°°ììŽ ìëŁë plateì ë©žê· ìêłŒ 1 ml ë©žê· ìë„Œ ë¶ìŁŒíìŹ íŹìë„Œ ë©žê· ìëĄ ìčšì¶ì쌰ìŒë©°, 10-mL syringeë„Œ í”íìŹ filteringí ë€ íìë íŹìì ëëë„Œ ë¶êŽêŽëêł(Synergy H1 Hybrid Multi-Mode microplate reader, BioTek, Winooski, VT, USA) OD600nm ëĄ 0.4 (106 cfu/ml)ëĄ ìĄ°ì íìë€.
6ìą
ë„ì PGPR íšì íì±ì êČìŠíììŒë©°, ì§ê· ìžíŹëČœ ìíŽ ë„ë „ì ì ì ìë chitinaseì cellulose íšì íì±êłŒ ìČ ìŽìš íĄì ë„ë „, ìžì° ê°ì©í íšì, ì§ì êł ì ë„ë „ì êČìŠíìë€. Chitinase íšì íì±ì êČìŠíêž° ìíŽ chitinolytic reaction mediaë„Œ ì ìĄ°íêł ì 5 g of chitin, 5 g of yeast extract, 0.7 g of KH2 PO4, 0.5 g of K2 HPO4, 0.3 g of MgSO4, 0.1 g of FeSO4, 0.1 g of NaClë„Œ 1 liter ìŠë„ìì ëŁêł pH 6.5-7.0ëĄ ìĄ°ì í ë€ì 18 g of agarë„Œ ìČšê°íìêł proteinase íì± ë„ë „ êČìŠì ìíŽ skim milk (10 g skim milk, 18 g agar per liter) ë°°ì§ë„Œ ìŹì©íìë€. ìČ ìŽìš íĄì ë„ë „ì êČìŠíêž° ìíŽ CAS agar (solution 1: 0.06 g of CAS, 50 ml of ddH2 O; solution 2: 2.7 mg FeCl3-6H2 O, 10 ml 10 mM HCl; solution 3: 73 mg hexadecyltrimethylammonium bromide, 40 ml ddH2 O; Blu dye: solution 1: 50 ml, solution 2: 9 ml, solution 3: 40 ml; mixture solution: MM9 salt solution 500 ml [15 g KH2 PO4, 25 g NaCl, 50 g NH4 Cl, 500 ml of ddH2 O]; 20% glucose stock 100 ml, NaOH solution [25 g NaOH per 150 ml ddH2 O]; casamino acid solution [casamino acid 3 g per 27 ml ddH2 O]; CAS agar: 100 ml MM9 salt solution, 32.24 g PIPES, 15 g agar, 30 ml casamino acid solution, 10 ml 20% glucose solution, 100 ml blue dye)ë„Œ ìŹì©íìë€. ìžì° ê°ì© ë„ë „ì êČìŠíêž° ìíŽ Pikovaskaya's ë°°ì§(10 g glucose, 0.5 g (NH4)2 SO4, 0.2 g KCl, 0.1 g MgSO4, 0.002 g MnSO4, 0.002 g FeSO4, 0.5 g yeast extract, 5 g Ca3(PO4), 20 g agar per liter, pH 6.8-7.0)ë„Œ ìŹì©íìë€(Kim ë±, 2019c). ì§ìêł ì ë„ë „ì êČìŠíêž° ìíŽ ìĄìì NFb ë°°ì§(5 g malic acid, 0.6 g K2 HPO4, 0.4 g KH2 PO4, 0.01 g MnSO4, 0.05 g MgSO4, 0.02 g NaCl, 0.002 g Na2 MoO4, 2 ml of bromothymol blue [0.5% in alcohol], 1.75 g of agar per liter, pH 6.6-7.0.)ë„Œ ìŹì©íìë€(Kim ë±, 2019c). IAA production assayë„Œ êČìŠíêł ì Salkowski reagent method (35% of HClO4 and 0.5 M of FeCl3)ë„Œ ìŹì©íìë€(PatelêłŒ Parmar, 2013).
íŽëč ì ì©ëŻžìëŹŒ 2ìą
ì ê°ê° tryptophan íšì ë ë°°ì§ì íšì ëì§ ìì PDK broth (10 g of potato dextrose broth, 2 g of Bacto peptone per liter)ì ì ìą
íìŹ 30°Cìì 14ìŒê° ë°°ìí ë€ì 5,000 rpm ììŹë¶ëŠŹêž°ëĄ 10ë¶ê° ë°ììíš í 1 mlì ìë±ìĄì test tubesì ìČšê°íìë€. ìŽ í Salkowski reagentì 2:1 ëčìšëĄ íŒí© í 25ë¶ìŽ êČœêłŒíìì ë ììì ëłíë„Œ êŽì°°íìë€(PatelêłŒ Parmar, 2013). Chitinase, siderophore, ì§ìêł ì ë„ë „, ê·žëŠŹêł protease íì± ë„ë „ì 28°Cìì ë°°ìíë©Žì 24ìê° ê°êČ©ìŒëĄ 3ìŒìŽ êČœêłŒí ìì êčì§ íìžíììŒë©°, ëš ì§ìêł ì ë„ë „ì ëìŒí ìšëìì ìŒìŁŒìŒê° ë°ìì êŽì°°íìë€.
pHì ë°ë„ž ìì„ ìĄ°ê±Ž ë° íŹì íì± ë„ë „ êČìŠ
ìëŹŒíì ì€ížë ì€ ì ê° ë° ê”°ì§í ë„ë „ìŽ êČìŠë ë°©ì ê· 2ìą
(Streptomyces sp. S8 and S. globisporus SP6C4) ê· ìŁŒì ìì„ íčì±ì íìžíêł ì pH 2, 5, 7, 7.2, 10 ìĄ°ê±Žìì ìì„ ìëë„Œ ë¶êŽêŽëêł(Synergy H1 Hybrid Multi-Mode microplate reader, BioTek) OD600nmìì 0.2ëĄ ìžĄì íììŒë©° íŹì íì± ë„ë „êłŒ ë°°ì§ ìĄ°ê±ŽëłëĄ ISP 2 media (yeast malt agar: yeast extract 4 g, malt extract 20 g, dextrose 4g, agar 20 g per liter), ISP 4 media (inorganic salt starch agar: CaCo3 2 g, K2 HPO3 1 g, MgSO4â§7H2 O 1 g, NaCl 1 g, (NH4)2 SO4 2 g, Trace salt solution [FeSO3â§7H2 O 0.1 g, MnCl2â§4 H2 O 0.1 g, ZnSO4â§7H2 O 0.1 g per 100 ml], ddH2 O 500-ml and adjusted pH 7.4), ISP 6 media (peptone yeast extract iron agar: peptone 15 g, proteose peptone 5 g, ferric ammonium citrate 0.5 g, dipotassium phosphate 1 g, sodium thiosulfate 0.08 g, yeast extract 1 g, agar 20 g per liter, adjust to pH 7.2), ISP 7 media (tyrosine agar: L-asparagine 1 g, L-tyrosine 0.5 g, dipotassium phosphate 0.5 g, magnesium sulphate·7H2 O 0.5 g, sodium chloride 0.5 g, sodium salt solution [ferrous sulphate·7H2 O 1.36 mg, copper chloride 2H2 O 0.027 mg, co-balt chloride 6H2 O 0.04 mg, sodium molybdate 2H2 O 0.025 mg, zinc chloride 0.02 mg, boric acid 2.85 mg, manganse chloride·4H2 O 1.8 mg, sodium tartarate 1.77 mg] agar 20 g per liter, adjusted pH 7.4). ê·žëŠŹêł MS (mannitol soya agar: mannitol 20 g, soya flour 20 g, agar 20 g per liter)ìì 24ìê° ê°êČ©ìŒëĄ 72ìê° ëì ë°°ìì í”íŽ íìžíìë€. íŹìíì±ì ê°ê°ì ë°°ì§ìì ë°©ì ê· ì êž°ê· ìŹ íì±êłŒ íŹìíì± ì ëë ìĄììŒëĄ íìžíìë€.
Phenotype assay ë„Œ ìŽì©í ì§ìì ì ížë êČìŠ
ëšìŒ ê· ìŁŒë„Œ ëììŒëĄ ìì„ ìê°ì ëšì¶ìí€êž° ìíŽ ìì„ì ìŽì§í ì ìë ì§ììì ì ëłíêł ì PM3B plate Biolog plate (Biolog, Hayward, CA, USA)ë„Œ ìŹì©íìë€. ê° plateìë OD600ìì 0.2ëĄ ìĄ°ì ë ë°©ì ê· íŹì ííìĄ 10 ”lë„Œ ë¶ìŁŒí í 28°Cìì 3ìŒê° ë°°ìíìë€. ë°°ììŽ ìëŁë plateì Redox dye mix Aë„Œ ê° wellì 10 ”l ë¶ìŁŒí í 37°Cìì 30ë¶ ê°êČ©ìŒëĄ ìì ëłíë„Œ êŽì°°íìêł ìŽí ì ë°ë ì§ììì 0.2% (final concentration)ìŒëĄ ì§ìììŽ êČ°íë basal media (KCl 0.05%, KH2 PO4 0.1%, MgSO4·7H2 O 0.05% and FeSO4·7H2 O 0.001%)ì ëëë„Œ 5 ÎŒ M, 10 ÎŒ M, 50 ÎŒ M, 5 mM, 10 mM, 50 mMëĄ ë€ë„ŽêČ ìČšê°íìŹ ê· ìŁŒì ìì„ ìëì ìí„ì 믞ìčëì§ êČìŠíìë€(Ripa ë±, 2009).
íìì ë° ì§ìì ìČšê°ì ë°ë„ž íŹì íì± ë„ë „ êČìŠ
íŹì íì± ë„ë „ êČìŠìì í볎ë íììêłŒ Biolog 3B plateë„Œ í”íŽ ìì„ì ìĄ°ì íë êČìŒëĄ ì ë°ë ì§ììì ìŽì©íìŹ ìĄì ë°°ì ì íŹì íì± ë„ë „ì êČìŠíìë€. íììêłŒ ì§ììì ê°ê° basal media (KCl 0.05%, KH2 PO4 0.1%, MgSO4·7H2 O 0.05%, FeSO4·7H2 O 0.001%)ì final concentration 0.2%ëĄ ìČšê°íììŒë©°, ëȘšë íììêłŒ ì§ììì 2% stockìŒëĄ ì ìĄ°íìŹ 0.2 ÎŒ m syringe filter í ìŹì©íìë€. ìŽí OD600 nm 0.2ëĄ ìĄ°ì ë íŹì ííìĄ 10 ”lë„Œ 1 mlì basal mediaì ì ìą
íìŹ 30°Cìì ì§íë°°ììŒëĄ 3ìŒê° ë°°ì í í믞êČœ(BX53 microscope, Olympus, Tokyo, Japan) 1,000ë°° ë°°ìšìì íŹìì íì± ì ëë„Œ êŽì°°íìë€.
êČ°êłŒ ë° êł ì°°
Streptomyces ì±ì„ì ììŁŒ ìëŹŒì êł”êžìì êŽì°°ëë êČìČëŒ ìì° ìĄ°ê±Žìì íčì ìì êł”êžìì ìíŽ í„ìë ì ìë€(Costa ë±, 2002; Reese ë±, 2018). ìëŹŒìì íčì íììêłŒ ì§ìì ìììë 믞ìëŹŒì ì±ì„êłŒ ë°ëë„Œ ì§ìíêł ìĄ°ì í ì ììŒë©°, ìžíŹ ì±ì„ì í„ììíŹ ëżë§ ìëëŒ ëŻžìëŹŒêłŒ ììŁŒì êž°ë„ì ìíž ìì©ì íŹêČ êž°ìŹíë€(Du ë±, 2021; Ducray ë±, 2019). ìŽë€ íí©ëŹŒì íì êž°ë„ì ê°ì§ ììČŽ íì± íí©ëŹŒ ë° ëšë°±ì§ì ìì°ì ìĄ°ì ìëĄ ìí ì ìííê±°ë íì©ëë€êł ëłŽêł ëìŽ ìë€(Costa ë±, 2002; Ricoì Preston, 2008; Voelker ì Altaba, 2001). ê·ž ììëĄ Kim ë±(2021)ì L-glutamic acid ê° ì§ì ë° ì§í ìëŹŒ ìĄ°ì§ ëȘšëìì í”ìŹ ë°í
늏ì ê· ìŁŒì ê°ìČŽê”°ì ìŹê±Žíêł , 믞ìëŹŒ ê”°ì§ ê”ŹìĄ°ë„Œ ìŹê”Źì±íë©° ëì 믞ìëŹŒ ë€ìì±ì ì ì§íìŹ ì”ìą
ì ìŒëĄ ìëŹŒ ëłììČŽì êł”êČ©ì ìë°©í ì ìë€êł ëłŽêł íìë€.
ë°©ì ê· ì ê· ìŁŒ íčìŽì±ìŽ ë§€ì° ëì íčì±ì ì§ëêł ììŒë©° 2ì°š ëìŹì°ëŹŒì íì±ìŽ ì€ìíë€(Mostafa, 1979). ëí ìíêł ëŽ ììĄŽì ìíŽ ììČŽ ìì°íë 2ì°š ëìŹ ëŹŒì§ ë¶íŽ ë„ë „ì ì§ëêł ììŒëŻëĄ, 1ì°š ëìŹë„Œ 충ë¶í ì§íí ìíìì 2ì°š ëìŹëĄ ì íìšì ëšìê°ì ëìŒ ì ìëëĄ ì ëíìŹìŒ íë€(Padilla ë±, 1991). ìŽì êž°ìŽ ë°°ì ìĄ°ê±Ž ëšêłëĄì ë°©ì ê· ì ìììê”Źì±ì ë°ëŒ ì ë°ë íìì, ì§ìì íŒí© ìĄ°ê±Žì ìŽì©íìŹ íŹì íì± ë„ë „êłŒ ìì„ ìë ìžĄì ì í”íìŹ ê· ìŁŒ ìì„ ìëì ììŽ ì°ìíë€êł 볎ìŹì§ë ë°°ì§ íŒí©íì ìŽì©íìŹ ì ë°íêł ì íìë€. ëí ë§ì§ë§ ëë ë°°ì ëšêłëĄì ë°°ììĄì ì ìą
ììŒëĄ íìŹ ëëë°°ìêž°ìì ìĄ°ì í ì ìë ì í© pH ìĄ°ê±Žì ì ë°íêł ì íìŹ ë°©ì ê· ì”ì í© ë°°ì ìĄ°ê±Žì í볎íêł ì íììŒë©°, ê· ìŁŒì íčì± ê”ŹëȘ
ì ìíŽ 6ìą
ë„ì PGPR íì± ë„ë „ì íšê» êČìŠíìë€.
PGPR íšì íì± êČìŠ êČ°êłŒ
ì€ëŹŒìììŒëĄ êž° í볎ë 2ìą
ì ë°©ì ê· ìž Streptomyces sp. S8êłŒ Streptomyces globisporus SP6C4ì ê°ê° ìëĄ ë€ë„ž êž°ìŁŒìì ë¶ëŠŹëìì§ë§ íì§ê· íì±ìŽ ëë€ë êł”í”ì ìž íčì§ì ì§ëêł ììë€(Cha ë±, 2016; Jeon ë±, 2019; Kim ë±, 2019a). ìŽì ëłž ì°ê”Źìì ë ê· ìŁŒì íčì±ì ê”ŹëȘ
íêł ì íì§ê· íì±ì ëíëŽë 2ìą
ìž chitinase enzyme productionêłŒ protease enzyme productionì íŹíšíìŹ ë€ìí íšì íì±ì êČìŠí ì ìë siderophore production, phosphate solubilization activity, nitrogenase enzyme production, IAA productionì íìžíìë€. ê·ž êČ°êłŒ S8 ê· ìŁŒë chitinase, protease, siderophore, phosphate solubilizationìì ëȘšë íì±ì ëíëìŒë©°, IAA íšì ìì± ë„ë „êłŒ ì§ìêł ì ë„ë „ì íìžëì§ ììë€(Table 1, Fig. 1). íì§ë§ SP6C4ë proteaseë„Œ ì ìží ëȘšë íšì íì± ë„ë „ìŽ ìë êČìŒëĄ íìžëìë€(Table 1, Fig. 1). ìŽ 6ìą
ë„ì PGPR íšì ë„ë „ êČìŠ êČ°êłŒ S8 ê· ìŁŒê° SP6C4 ëłŽë€ ë€ìí íì±ì ëíëŽë êČìŒëĄ íìžëìë€.
Fig. 1.
Plant growth-promoting rhizobacteria activity of Streptomyces sp. S8 and S. globisporus SP6C4. The spore stock solution (10 ”l) was added to each enzyme assay medium that contained an 8 mm diameter filter paper disk. (A) Chitinase enzyme production (left), Protease enzyme production (right). (B) Siderophore production, phosphate-solubilization activity, nitrogenase enzyme production, and indole-3-acetic acid production. The upper panel in each figure showed S8 and the lower panel presented SP6C4 strain.

pHì ë°ë„ž ìì„ ìĄ°ê±Ž ë° íŹì íì± ë„ë „ êČìŠ êČ°êłŒ
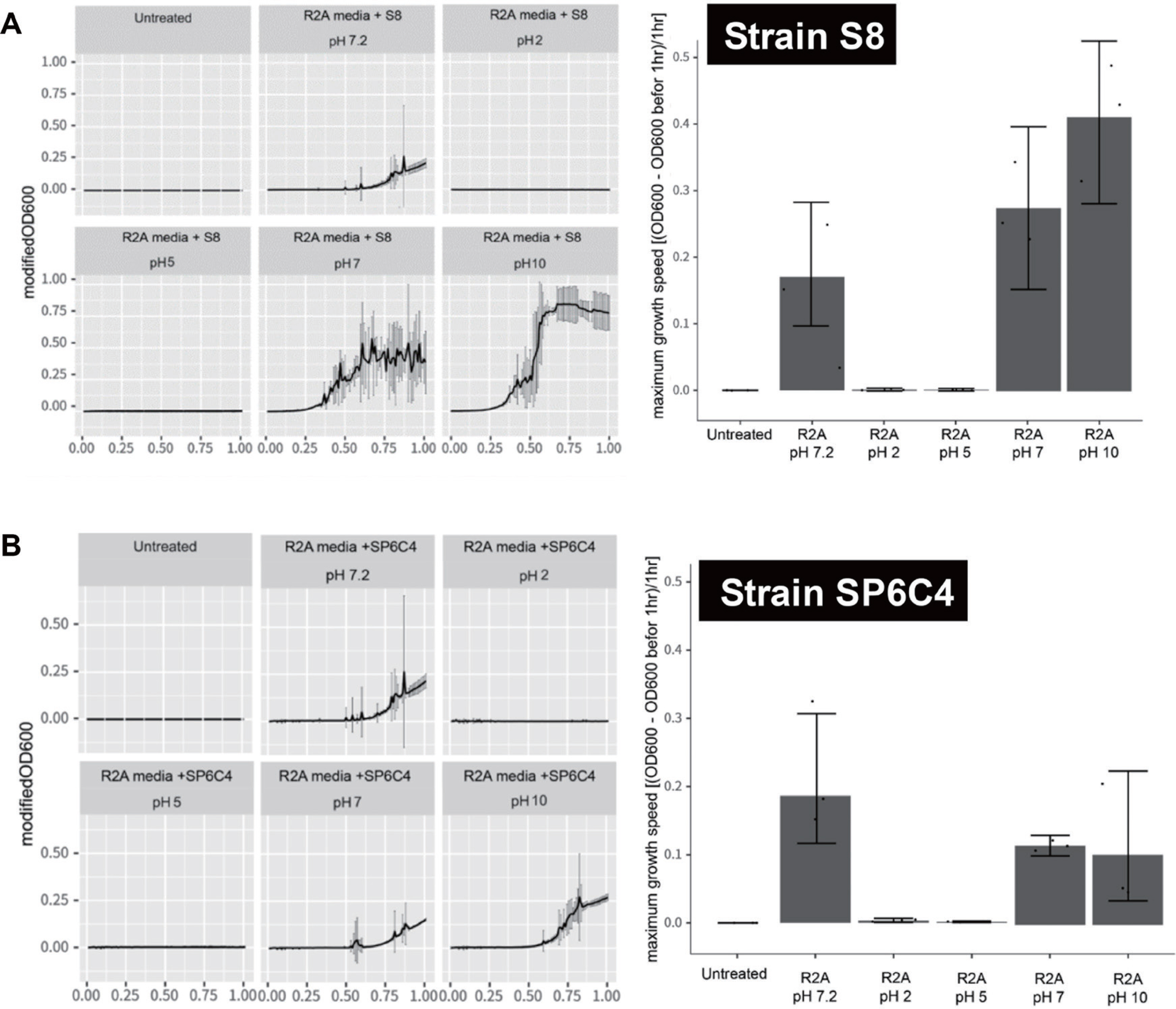
ëë ë°°ì ì ìŽêž° ìì„ë„ ì ìŽì§í ì ìë ì í© pHë„Œ êČìŠíêł ì ì°ì± ìĄ°ê±Žìž pH 2ì pH 5, ì€ì± ìĄ°ê±Žìž pH 7ì pH 7.2, ê·žëŠŹêł ìŒ êž° ìĄ°ê±Žìž pH 10ì ìŽì©íìŹ ë°©ì ê· ì ìì„ë„ êłŒ ëčìì„ë„ (êž°ìžêž°ë„Œ ìŽì©íìŹ ìê°ëč ì”ë ìì„ë„ ) ìžĄì íìë€. ê·ž êČ°êłŒ S8ê· ìŁŒë pH 7êłŒ pH 10ìì ìì„ìŽ ê°ë„íšì 볎ììŒë©° ìŒêž° ìĄ°ê±Žìž pH 10ìì ê°ì„ ëč ë„ž ìì„ë„ ì íìží ì ììêł , SP6C4ë ì€ì± ìĄ°ê±Žìž pH 7, pH 7.2 ê·žëŠŹêł ìŒêž° ìĄ°ê±Žìž pH 10ìì ìì„ë„ ì ëíëì§ë§ ì€ì±ìž pH 7.2ìì ê°ì„ ëì ìì„ë„ ì ëíëŽë êČìŒëĄ íìžëìë€(Fig. 2).
Fig. 2.
Variation with pH in bacterial growth. The spore stock (10 ”l) of the suspension was added to each well of a 96-well plate. The plates were incubated, and the optical density was measured with orbital shaking using a Synergy H1 Hybrid Multi-Mode microplate reader for a period of 24 hr at a temperature of 28°C. (A) S8 strain. (B) SP6C4 strain. The left panel showed bacterial growth in each pH condition. The right panel presented the maximum growth speed of the strain.
íŹì íì± ë„ë „ êČìŠì ìŹì©í 5ìą
ë„ì ë°°ì§ìì 24ìê° ê°êČ©ìŒëĄ 72ìê°êčì§ íìží êČ°êłŒ S8êłŒ SP6C4 ëȘšë 24ìê°ìì ISP7ìì ê°ì„ ìœí íŹì íì± ë„ë „ì ëíëìë€(Fig. 3A). 48ìê°ìŽ êČœêłŒíìì ë S8ì ISP2ì ISP6 ê·žëŠŹêł MSìì ëč ë„ž íŹì íì± ë„ë „ìŽ íìžëììŒë©° ìŽë SP6C4ë ì ìŹí êČ°êłŒëĄ íìžëìë€(Fig. 3B). 72ìê° êČœêłŒ í 5ê°ì§ ë°°ì§ìì ëȘšë íŹì íì±ìŽ íìžëììŒë©° S8ìŽ SP6C4ëłŽë€ ìŽêž° íŹì íì± ë„ë „ìŽ ì°ìíë©°, ë ê· ìŁŒ ëȘšë êł”í”ì ìŒëĄ yeastì mannitolì ìŁŒ í ìììŒëĄ íšì ë ë°°ì§ìì íŹì íì±ìŽ ì ëëë êČìŒëĄ íìžëìë€(Fig. 3C).
Fig. 3.
The emergence of phenotypic in colonies of Streptomyces sp. S8 and S. globisporus SP6C4. Five different media, ISP2, ISP4, ISP6, ISP7, and MS (from left to right) were tested. The plates were incubated at 28°C for 72 hr, and observations were made at three different time points: (A) 24 hr after incubation, (B) 48 hr after incubation, and (C) 72 hr after incubation. The upper panel showed S8 and the lower panel presented SP6C4 strain.

Phenotype assayë„Œ ìŽì©í ì§ìì ì ížë êČìŠ êČ°êłŒ
íìììŒëĄì 2ê· ìŁŒ ëȘšë ì ì© ê°ë„í yeastì mannitolì ì ë°í ë€ì ì§ììì ì ë°íêł ì Biolog 3B plateë„Œ ì€íì ìŹì©íìë€. ê·ž êČ°êłŒ S8ì ëšìŒ ì§ìììŒëĄ ammoniaì L-cysteine ìì ìì„ìŽ ìŽëŁšìŽì§ êČì íìžíììŒë©° SP6C4ë L-cysteine, L-glutamic acid, L-prolineìŒëĄ íìžëìë€(Fig. 4). ê°ê° ì ë°ë ì§ììì ëëì ë°ëŒ ìŹì©íìŹ ìì„ë„ ì íìží êČ°êłŒ S8ì L-cysteineìŽ 50 mM ëëëĄ ìČšê°ëìì ë, ê·žëŠŹêł SP6C4ë L-glutamic acid ìČšê°ëìì ë ì”êł ëëìž 50 mMì ì ìžíêł ìì„ìŽ ëȘšë ìŽì§ëë êČì íìžíìë€(Fig. 5).
Fig. 4.
Biolog phenotype array for nitrogen utilization by PM3B plate. The spore stock was inoculated onto PM3B plates. A re-dox dye mix A was added to the plate, and the plate was further incubated at 37°C. Numbers (1-12) and alphabet (A-H) repre-sented well position and each position contained different nutrient sources.
Fig. 5.
Effect of differences in nitrogen sources to bacterial growth. The nitrogen stock solution was added to basal media, followed by the addition of bacterial spore stock standardized to an optical density of 0.2 at 600 nm. The mixture was then incubated with shaking at 28°C for a duration of 50 hr (x-axis).

íìì ë° ì§ìì ìČšê°ì ë°ë„ž íŹì íì± ë„ë „ êČìŠ êČ°êłŒ
ê°ê°ì ì ë°ë íìì 2ìą
êłŒ ì§ìì 3ìą
ì íŹíšíìŹ ë°©ì ê· íŹì íì±êłŒ 2ì°š ëìŹì°ëŹŒ íì± êłŒì ì êŽìŹíë€ êł ëłŽêł ë L-asparagine, L-valine, peptoneêłŒ starchë„Œ ìŽì©íìŹ ìĄì ë°°ì ìĄ°ê±Žììì íŹì íì± ë„ë „ì êČìŠíìë€(Kalaiyarasi ë±, 2020; Mostafa, 1979). ê·ž êČ°êłŒ S8ë íìì ì€ starchì mannitolìì íŹì íì±ìŽ ìŽëŁšìŽì§ì§ ìììŒë©° aggregationëë êČìŽ êŽì°°ëìêł , yeast extractìì íŹì íì±ìŽ íìžëìë€(Fig. 6). ë°ë©Ž ì§ìì 6ìą
ì ëíŽìë ëȘšë ìì±í íŹì íì±ìŽ íìžëìë€(Table 2, Fig. 6). ë€ë„ž ë°©ì ê· ìž SP6C4ììë íìì ì€ yeast extractììë§ íŹì íì±ìŽ ìŽëŁšìŽìĄìŒë©° ì§ììì êČœì° L-glutamic acidêłŒ L-prolineììë§ íŹì íì±ìŽ êŽì°°ëìë€(Fig. 7). íčìŽì ìŒëĄ peptoneìŽ ìČšê°ë êČœì° SP6C4ìì êž°ê· ìŹê° ìì±íêČ íì±ë êČì íìžíìë€(Table 2, Fig. 7).
Fig. 6.
Spore chain formation with different carbon and nitrogen sources by Streptomyces sp. S8. Six different types of nitrogen sources and three types of carbon sources were added to basal media at a final concentration of 0.2%. The sporulation was visualized under a 1,000Ă microscope (scale bars=10 ÎŒm). (A) L-asparagine. (B) L-cysteine. (C) L-glutamic acid. (D) L-proline. (E) L-valine. (F) Peptone. (G) Starch. (H) Mannitol. (I) Yeast extract.

Fig. 7.
Spore chain formation with different carbon and nitrogen sources by Streptomyces globisporus SP6C4. Basal media was supple-mented with six different nitrogen sources and three different carbon sources, each at a final concentration of 0.2%. The sporulation was observed under a 1,000Ă microscope (scale bars=10 ÎŒm). (A) L-asparagine. (B) L-cysteine. (C) L-glutamic acid. (D) L-proline. (E) L-valine. (F) Peptone. (G) Starch. (H) Mannitol. (I) Yeast extract.

Table 2.
Validation of spore formation ability under microsco-py as affected by nitrogen and carbon sources
| Type | Sources | Sporulation a | |
|---|---|---|---|
| SP6C4 | S8 | ||
| Nitrogen | L-asparagine | - | Y |
| L-cysteine | - | Y | |
| L-glutamic acid | Y | Y | |
| L-proline | Y | Y | |
| L-Valine | - | Y | |
| Peptone | -ah | Y | |
| Carbon | Starch | - | - |
| Yeast extract | - | Y | |
ëłž ì°ê”Źììë êž° í볎ë ì ì© ë°©ì ê· (Streptomyces)ì 2ìą
ì ìëŹŒ ì ë ìčíêČœ 믞ìëŹŒ ì ì ëĄ ê°ë°íšì ììŽ ì ì”ì±ì ì êł”íêł ì ê·ž íčì±ì ê”ŹëȘ
íêł ì í© ë°°ì ìĄ°ê±Žì í볎íë €êł íìë€. ê·ž êČ°êłŒ S8êłŒ SP6C4 ë°©ì ê· ì PGPRì íšë„ìŽ ìëĄ ììŽí íčì±ì íì©íìŹ íŒí© ë°°ì íëłŽê”°ìŒëĄì ê·ž êž°ë„ìŽ ë ëêČ íê°ë êČìŒëĄ ìê°ëë€. íìììŒëĄ yeast extract ëë mannitol, ì§ìììŒëĄë L-cysteinì êž°ëłž ìí„ììŒëĄ ìŹì©í êČœì°, S8êłŒ SP6C4ì êł”ë°°ìì íšìšì ìž êČ°êłŒë„Œ ëì¶í êČìŒëĄ ìê°ëë€. íì§ë§ ë ë°©ì ê· ì ììĄ ì”ì ì°ëìĄ°ê±ŽìŽ S8ì pH 10, SP6C4ë pH 7.2 ììŽíìŹ ìŽì ëí íì ì°ê”Ź ìíìŽ ìê”Źëë€. ëí 믞ìëŹŒ ì ì ëĄì ê°ë°íšì ììŽ íŹìì ííëĄ ìĄŽìŹí êČœì° ì„êž°ëłŽêŽì ìì ì±ìŽ ëìì§êł 볎êŽìŽ ì©ìŽíëŻëĄ, ëłž ì°ê”Źë„Œ í”íŽ ì ë°ë íìì·ì§ììì í”íìŹ íŹì íì±ì ìŽì§í ì ììŒëŻëĄ 믞ìëŹŒ ì ì ê°ë°ì êž°ìŽì ìëŁë„Œ ë§ë šíìë€.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






