



Introduction
Maize (Zea mays L.) is amongst the widely grown staple crops in the world, ranking third after wheat (Triticum aestivum L.) and rice (Oryza sativa) (Reeves et al., 2016). Maize is a major staple food and an important feed crop for livestock (Berger et al., 2020). Maize is a rich source of nutrients, including carbohydrates (84.0%), proteins (10.9%), fats (4.5%), and minerals (1.3%), which makes this crop ideal for food and feed (Romney et al., 2003). The crop is also important in processing of food products, tanning, biofuel, and cosmetic industries including starch, sweeteners, oil, beverages, glue and etc. (Ranum et al., 2014).
In South Africa, the Eastern Cape Province has a high proportion of households (20%) involved in agricultural activities, primarily smallholder farmers who grow maize for subsistence. These farmers often use low-input production practices, resulting in low yields. Yield losses can be attributed to various abiotic and biotic factors, including water, climate, soil status, and plant diseases, all of which can ultimately affect overall crop performance. Fungal foliar diseases are a significant challenge in maize production (Berger et al., 2020), and one of the most prominent diseases in the Eastern Cape is northern corn leaf blight (NCLB) caused by Exserohilum turcicum (Wang et al., 2014), which has deleterious effects on maize production globally (Chandrashekara et al., 2014).
The E. turcicum pathogen is known to survive under moderate temperatures, long heavy dew periods (Welz and Geiger, 2000), humid weather, and frequent rainfall (Rashid et al., 2020). Infection is promoted through conidia germination and sporulation when free water is present on the leaf surface for 6-18 hr at temperatures between 18-27°C (Hilu and Hooker, 1964). Subsequently, lesion-induced spores rapidly disperse the disease. The pathogen can overwinter in soil and plant residues, allowing it to survive unfavorable conditions. During favourable conditions, E. turcicum is dispersed by wind and rain splash (Hooda et al., 2017).
The impact of NCLB on host plants depends on their developmental stage. Wathaneeyawech et al. (2015) reported that maize plants infected by E. turcicum at the vegetative stage exhibit severe yield reduction of up to 70%. Yield losses at the reproductive stage can amount to 40-70% (Levy and Pataky, 1992). Maize grain yield losses incurred from the disease may be up to 44% from disease severities of 52-100% at the full dent stage (Bowen and Pederson, 1988). Therefore, after maximum yield biomass accumulation, the pathogen has a limited effect on the crop.
For the past five years, most maize cropping projects in O.R. Tambo district have faced a significant challenge of high infestation by NCLB. Smallholder farmers, who are often impacted by this disease, lack knowledge regarding disease management and the types of resistant maize genotypes available to combat NCLB. While using resistant genotypes against NCLB is prudent, information on NCLB-resistant varieties in South Africa is lacking, including their availability. Additionally, there is a paucity of research and knowledge on the severity, impact, and management strategies of NCLB fungal foliar disease in smallholder farms in the Eastern Cape. Therefore, conducting research on NCLB fungal foliar disease within the context of smallholder maize production in the Eastern Cape is critical. This study aimed to assess NCLB disease severity and its impact on the growth and yield of various maize genotypes, focusing on demonstration plots in the O.R. Tambo district.
Materials and Methods
Site description
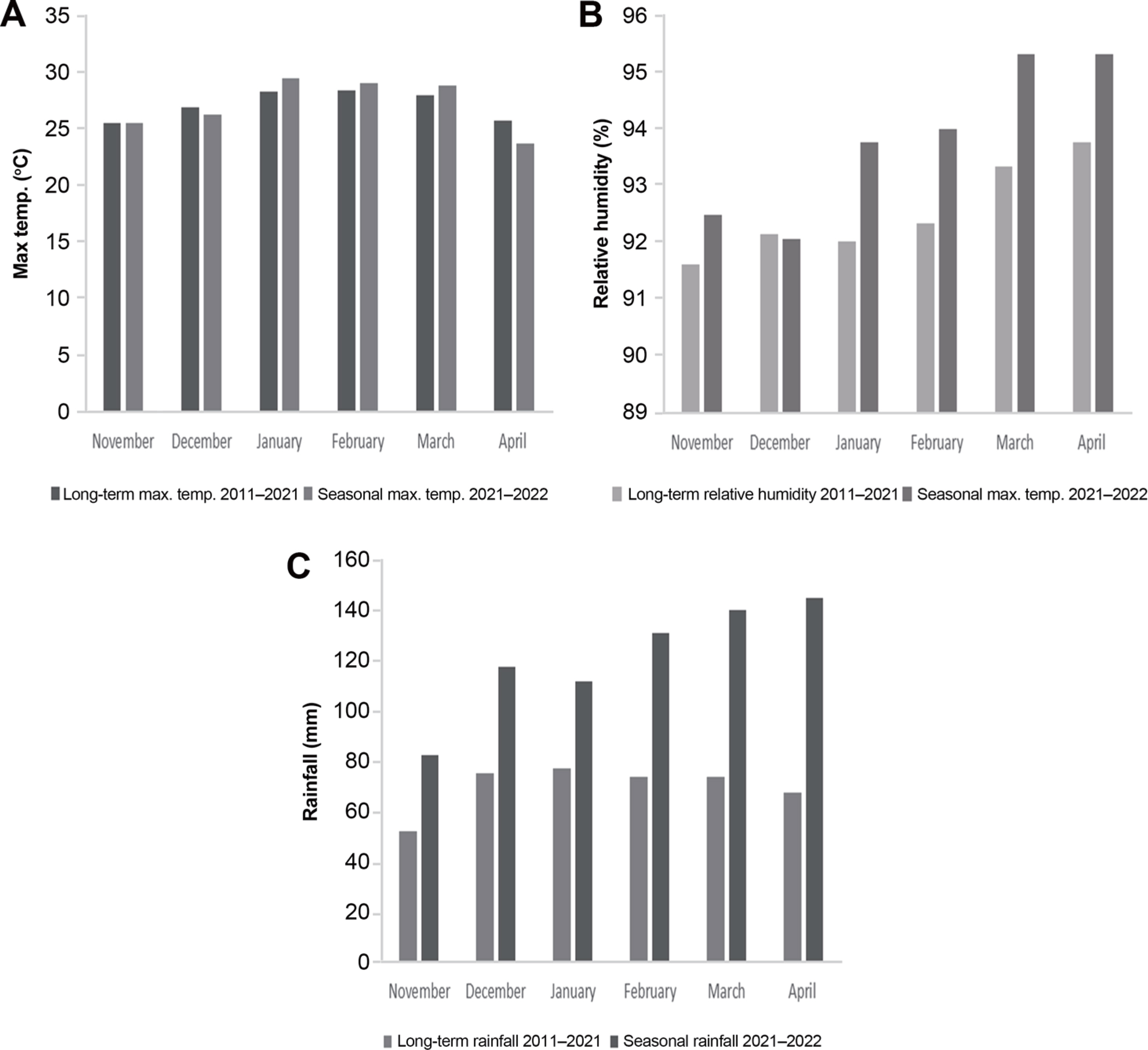
The current experiment was conducted at Njiveni (31°34′5,79”S and 29°8′33,48”E) maize field located in Ngqeleni municipality, O.R. Tambo district in 2021/2022 growing season under rainfed conditions. Seasonal climatic data was recorded, and long-term climatic data was attained from Agricultural Research Council (ARC), South Africa (Fig. 1). The area experiences an average of 800 mm rainfall per annum and the average daily temperatures ranging between 25°C and 30°C.
Experimental design and agronomic management
A randomized complete block design with three replications was used for this study. Ten maize cultivars were sourced from three seed companies. PAN 5R-582R, PAN 4R-528R, PAN 5R-890R, PAN 4R-728BR, PAN 3R-724BR, PAN 5R-854R, and PAN R5R-590R were sourced from Pannar; DKC 74-74BR and DKC80-40BR from Monsanto; and P1788B from Pioneer seed company. Maize seeds were planted on a plot of 0.0025 ha (25 m2) with an intra-row spacing of 25 cm and an inter-row spacing of 75 cm. Two maize seeds were planted per planting station and thinned to a plant population of approximately 53,333 plants/ha. Ten plants were randomly selected from the two middle rows of each plot and tagged for disease assessment. A recommended rate of NPK fertilizer based on soil nutrient analysis (Table 1) was applied at planting. Limestone Ammonium Nitrate (LAN 28%) was side-dressed six weeks after planting. Cypermethrin was used to control maize insect pests at a rate of 300 ml/ha, and no fungicide was applied.
Maize growth and yield attributes
Plant height and leaf area: After reaching the milk stage, maize plant height, and leaf area were measured using 10 plants from the two middle rows of each plot. Plant height was recorded up to the base of the tassel, along the above-ground stem.
Leaf length (L) and leaf width (W) of each live leaf was measured and leaf area was calculated according to Dwyer and Stewart (1986) using Eq. (1):
Where L=leaf length, W=maximum leaf width, and A=is a constant of 0.75. The average leaf area of each plant was recorded.
Maize yield components: At harvest maturity, the maize crop was harvested and the unshelled cobs were weighed using a weighing scale. Thereafter, the cobs were hand shelled and fresh grain weight was measured. The grain moisture content was then measured using grain moister meter (LDS-1H/1S, Zhejiang Top Cloud-agri Technology Co Ltd., Hangzhou, Zhejiang, China) and air dried down to 12.5%. Adjusted grain yield was calculated according to Eq. (2) (Aliyi et al., 2018):
Where FW=field weight; AMC=actual moisture content; RDW=recommended dry weight (100-12.5)=87.5; 0.8=Given shelling percentage.
NCLB disease assessment
O.R. Tambo district in the Eastern Cape has experienced several planting seasons with maize plants showing signs of NCLB prior to this research. The environmental and climatic conditions in the district are conducive to the thriving of the NCLB pathogen. Therefore, maize plants were naturally infested with NCLB after planting.
Disease severity was assessed by recording the percentage of leaf area infected at the silk dry stage. Infected leaves were scored based on nine broad categories according to Nutter et al. (1991). These severity scores were then converted to percentage disease index (PDI) using the equation described by Wheeler (1969) in Eq. (3):
The PDI values were then arcsine transformed and subjected for Analysis of Variance.
Maize genotypes were classified as resistant or susceptible to the NCLB pathogen based on the criteria by Abdelsalam et al. (2022).
Statistical analysis
Analysis of variance (ANOVA) was performed using the SAS version 9.4 statistical package. Significant differences among means were separated using the least significant difference test at a 5% significance level. Pearson's correlation coefficients were calculated using MS Excel 2019 to determine relationships between maize parameters and tested for significance at P≤0.05.
Results
Plant height
Significant differences (P<0.05) in plant height were observed among maize hybrids. The hybrid DKC74-74BR exhibited the tallest average plant height of 207.0 cm, whereas PAN 5R-590R had the shortest average plant height of 185.3 cm. The average plant height of all other hybrids did not differ significantly (Table 2).
Table 2.
Growth and yield parameters of various maize genotypes
Table 3.
Percentage diseases incidence (PDI) among various maize genotypes
Leaf area
The leaf area of maize hybrids differed significantly (P<0.05) in this study. DKC74-74BR recorded the highest leaf area of 603.9 cm2 while hybrid PAN 5R-590R attained the lowest leaf area of 488.0 cm2 as shown in Table 2. It is interesting to note that hybrid PAN 5R-890R and DKC80-40BR are statistically similar with leaf areas of 518.4 cm2 and 518.0 cm2, respectively.
Cob weight
A significant difference (P<0.05) was observed in cob size (weight) between the maize hybrids. The largest cob size was observed in hybrid PAN 5R-582R, with a weight of 112.7 g/cob, as shown in Table 2. Hybrids PAN 5R-890R and PAN 3R-724BR had smaller cob weights in the current study, weighing 70.7 g/cob and 77.3 g/cob, respectively. These two hybrids were statistically similar. It is also worth noting that hybrids DKC74-74BR, DKC80-40BR, and PAN 4R-728BR had no significant differences between them, with cob sizes of 112.0 g/cob, 109.7 g/cob, and 109.7 g/cob, respectively. Similarly, hybrids PAN 5R-590R, PAN 5R-854R, and P1788BR did not differ significantly from each other, with cob sizes of 96.7 g/cob, 92.0 g/cob, and 90.7 g/cob, respectively (Table 2).
Grain fresh weight
There were significant differences (P<0.05) in the grain weights of the genotypes used in this research. The highest grain weight was observed in genotype PAN 5R-590R, which weighed 11,783.3 kg/ha. The smallest grain size in terms of fresh weight was observed in genotype DKC80-40BR, which weighed 7,188.9 kg/ha. Notably, hybrids PAN 5R-854R (7,511.1 kg/ha) and P1788BR (7,514.4 kg/ha) were statistically similar. Genotypes PAN 4R-728BR, PAN 5R-582R, and PAN 3R-724BR had statistically similar grain sizes, weighing 7,868.9 kg/ha, 7,955.6 kg/ha, and 8,041.1 kg/ha, respectively (Table 2).
Grain yield
Maize genotypes used in this study significantly affected grain yield (P<0.05). Greater grain yield was obtained by hybrid DKC 74-74 BR (8,333.3 kg/ha). Hybrids that performed poorly in terms of their grain yield were P1788BR (5,631.5 kg/ha), PAN 5R-854R (5,594.2 kg/ha), and DKC80-40BR (5,062.6 kg/ha) (Table 2). Hybrids PAN 4R-728BR and PAN 3R-724BR did not show any statistically significant differences in their grain yield. Similarly, hybrids PAN 4R-528R, PAN 5R-890R, and PAN 5R-590R also had no statistically significant differences in their grain yield.
Percentage disease index
Maize hybrids showed significant (P<0.05) variation in susceptibility to NCLB, as indicated by the PDI presented in Table 2. PAN 4R-728BR exhibited the highest susceptibility to NCLB at 70.9%, which was statistically similar to PAN 3R-724BR (68.7%) and PAN 4R-528R (67.9%). Hybrids DKC80-40BR and PAN 5R-582R had statistically similar disease incidences of 22.1% and 26.1%, respectively.
Among the 10 genotypes used in this study, four (PAN 4R-528R, PAN 4R-728BR, PAN 3R-724BR, P1788BR) were classified as susceptible, five (DKC74-74BR, PAN 5R-582R, PAN 5R-890R, PAN 5R-854R, PAN 5R-590R) were moderately susceptible, and one (DKC80-40BR) was classified as moderately resistant.
Correlation between PDI, vegetative growth, and yield attributes
A moderate and positive correlation (0.572) was found between leaf area and plant height, which was highly significant (P<0.01). Table 4 shows that all other plant growth and yield parameters had a weak and non-significant correlation. However, it is important to note that yield and fresh grain weight had a strong positive correlation of 0.739, although it was not statistically significant.
Table 4.
Correlation between maize growth, yield parameters and NCLB disease index
Discussion
Plant growth and zevelopment
This research aimed to assess the severity of NCLB disease on the growth and yield of various maize genotypes. Significant differences (P<0.05) were observed between hybrids in terms of growth parameters (plant height and leaf area) and yield attributes (cob weight, grain fresh weight, and adjusted yield), indicating the effect of genetic composition on the response to NCLB. These results align with the findings of Tahir et al. (2008), who also reported that genotype significantly impacted growth parameters in maize.
The genotype DKC74-74BR exhibited the highest growth and development performance, as indicated by both plant height and leaf area. Furthermore, this hybrid had the highest yield and the second-highest cob weight. These findings are consistent with those of Mtyobile (2021), who reported that higher cob production is positively associated with grain yield.
Plants that anchored high achieved high grain and cob yields by optimizing exposure to solar radiation, which promotes efficient photosynthesis. Tall genotypes also had wider leaves, increasing the surface area available for solar radiation absorption during photosynthesis. Optimal photosynthesis promotes the accumulation of greater photosynthates, which are necessary for grain development and filling. Photosynthates are translocated from sources, such as the stem and leaves, to sinks, including grains and cobs.
PDI
Significant differences in maize PDI scores were observed among different genotypes in this research, which is consistent with findings reported by Aliyi et al. (2018) and Abebe et al. (2008). These differences in PDI scores may be attributed to varying genetic potentials for disease susceptibility or resistance among different hybrid varieties (Aliyi et al., 2018). According to Kumar et al. (2011), resistance to NCLB disease in most maize genotypes is determined quantitatively, and this approach has been primarily used for disease control. Similarly, genetic diversity was used in our current research as a management strategy against NCLB disease in maize. Planting resistant genotypes is the most effective, economically feasible, and environmentally safe method for sustainable disease control.
Genotype DKC74-74BR showed the highest growth and development performance, yield, and the third lowest disease incidence. Despite being moderately susceptible to E. turcicum, this genotype had the largest leaf area. Moderately susceptible varieties (DKC74-74BR, PAN 5R-582R, PAN 5R-890R, PAN 5R-854R, and PAN 5R-590R) performed better than the susceptible varieties (PAN 4R-528R, PAN 4R-728BR, PAN 3R-724BR, and P1788BR). Furthermore, the moderately susceptible varieties performed even better than the moderately resistant genotype. These genotypes also showed relatively greater performance in terms of leaf area and plant height, which may have contributed to efficient photosynthesis (Walters and Bingham, 2007). Therefore, it can be inferred that moderate susceptibility to the disease did not significantly affect plant performance in this study, potentially remaining below the economic injury level at which the pathogen would cause significant damage to the crop.
DK80-40BR had the least disease incidence and was classified as moderately resistant, but it also had the lowest fresh grain mass and yield. The only notable trait in which this genotype outperformed the others was cob weight. These findings suggest that developing resistance against NCLB may come at the expense of plant growth and yield performance, consistent with the phenomenon of tradeoffs between different traits (Dwivedi et al., 2020, 2021). In the current study, resistance developed in DK80-40BR even though most of its plant performance traits were negatively impacted, except for cob weight.
Correlations
There was a significant positive correlation between leaf area and plant height, with both traits playing an important role in light exposure and interception by leaves. Plant height increased with an increase in leaf area. The current study established that plant height, leaf area, and cob weight all had a positive correlation with grain yield. This is consistent with the findings of Xu et al. (2022), who reported a positive relationship between grain yield per unit mass and leaf area. Leaf area, stem, and root dynamics in maize all contribute to grain yield on a mass basis, as they are responsible for the acquisition of resources necessary for photosynthesis. Subsequently, this results in the accumulation of photoassimilates that contribute to grain filling.
It is noteworthy that the PDI had a negative correlation with plant height, leaf area, cob weight, and grain yield. Aliyi et al. (2018) and Debela et al. (2017) reported similar negative associations between disease incidence/severity and maize yield and yield components in different genotypes. Conversely, previous research has reported a positive correlation between NCLB resistance and plant height (Abdelsalam, 2022; Altaf et al., 2016; Razzaq et al., 2019).
In the current research, we observed a decrease in plant height, leaf area, and cob weight with an increase in PDI, which subsequently had a negative impact on maize yield. Veerabhadraswamy et al. (2014) also reported that disease severity has a profound effect on yield losses in maize. Bowen and Pedersen (1988) reported that for every 10% increase in disease severity, there is a corresponding yield loss of 2-8%. This may be attributed to the plant's allocation of resources and nutrients to strengthen defense mechanisms under biotic stress, which can have a detrimental effect on plant growth and reproduction traits (Herms and Matsson, 1992).
In a previous study, Abebe et al. (2008) found a significant negative association between disease severity and seed weight and yield. However, in the present study, we observed contrasting significance statuses for disease severity in its correlation with seed weight and yield. These discrepancies may be attributed to variations in disease severity in the studied regions, regional effects on the severity of the disease, and differences in the genotypes used.
The objective of this study was to assess the severity of NCLB disease on the growth and yield parameters of various maize genotypes. The results of this study indicate significant differences between hybrids in terms of growth parameters, yield attributes, and PDI. High PDI was found on maize hybrids PAN 4R-528R, PAN 4R-728BR, PAN 3R-724BR, and P1788BR, indicating that these hybrids were susceptible to NCLB. Hybrids DKC74-74BR, PAN 5R-582R, PAN 5R-890R, PAN 5R-854R, and PAN 5R-590R were moderately susceptible, while DKC80-40BR was moderately resistant. DKC80-40BR also had a greater cob weight, while DKC74-74BR was superior in all other plant growth and yield components. Although NCLB has detrimental effects on maize crop performance, it was evident from our research that crop performance and disease resistance had tradeoffs. Breeders may select hybrid DKC80-40BR to breed for resistance to NCLB in order to reduce maize susceptibility to the disease. However, DKC74-74BR may be selected to enhance crop performance such as optimum plant growth, development, and yield. Further selection of these genotypes for resistance against NCLB is still required to develop a sustainable and economical way of managing NCLB pathogens. The experiment needs to be repeated in different environments and under different disease management strategies using the hybrids employed in the current research.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






