



Bacterial wilt of tomato plants has been known as a devastating soil-borne disease caused by Ralstonia solanacearum species complex, and this bacterium invades roots and colonises vascular tissues of a broad range of economically important crops, including peanut, potato and geranium (Choi et al., 2022; Muthoni et al., 2014; Swanson et al., 2005). Chemical and biological controls have been suggested for managing the tomato bacterial wilt. The tomato bacterial wilt was efficiently reduced by various chemical treatments: copper-associated pesticides, streptomycin-validamycin A antibiotics mixture, combined hydrogen peroxide-nitric oxide, plant essential oils such as clove oil and palmarosa oil (Hong et al., 2013; Lee et al., 2012; Pradhanang et al., 2003). Bacterial and fungal species originating from a diverse plant rhizosphere also ameliorated bacterial wilt by enhancing plant immunity in tomato plants (Jogaiah et al., 2013; Kurabachew and Wydra, 2013; Tan et al., 2013). However, it is still necessary to investigate how to integrate different controlling methods into eco-friendly pest management programs during tomato production.
Changes in plant nutrient conditions have affected the disease severities of various crops such as tomato plants (Dordas, 2008; Tripathi et al., 2022). Foliar application of potassium nitrate (KNO3) reduced lesion size on Alternaria solani-inoculated tomato leaves (Blachinski et al., 1996). However, increasing nitrate and ammonium nitrogen nutrition led to increased disease susceptibility of tomato plants to bacterial speck and bacterial wilt caused by Pseudomonas syringae pv. tomato and R. solanacearum, respectively (Ding et al., 2021). Interestingly, ammonium-nitrogen increased tomato crown and root rot by Fusarium oxysporum f. sp. radicis-lycopersici infection, whilst nitrate-nitrogen ameliorated the disease severity (Duffy and Dé fago, 1999). CaCl2 supplement in nutrient solution decreased disease severity of bacterial canker by Clavibacter michiganense subsp. michiganense and bacterial wilt by R. solanacearum in tomato plants (Berry et al., 1988; Yamazaki and Hoshina, 1995). Enhanced plant immunity, such as elevated H2 O2 levels and increased peroxidase and polyphenol oxidase activities, was involved in the CaCl2-mediated suppression of tomato bacterial wilt (Jiang et al., 2013). Micronutrients such as boron, manganese and zinc could also altered disease development of several tomato plants. Boron levels in the nutrient solution was negatively correlated with tomato bacterial wilt severity (Jiang et al., 2016). Increased MnSO4 concentration in the nutrient solution reduced tomato black leaf mold caused by Pseudocercospora fuligena (Heine et al., 2011). Powdery mildew of tomato plants by a natural inoculation was suppressed by foliar spraying manganese (Mn-EDTA) and zinc (Zn-EDTA), each alone or in combination (Bakeer et al., 2012). Early blight in tomato plants inoculated with Alternaria solani was ameliorated by ZnSO4 supplement (Awan et al., 2019). Interestingly, zinc (Zn-EDTA)-amended hydroponic nutrient solution alone could not reduce tomato Fusarium crown and root rot by F. oxysporum f. sp. radicis-lycopersici, but the zinc improved biocontrol activity by Pseudomonas fluorescens (Duffy and Dé fago, 1997). Micronutrient iron also has could alter disease severity in tomato plants. Foliar spraying Fe-EDTA reduced powdery mildew of tomato plants (Bakeer et al., 2012). Limited iron supplement in the hydroponic cultures accelerated wilting of a resistant tomato cultivar against Verticillium dahlia infection, indicating the critical role of iron in plant immunity (Macur et al., 1991). However, information on the micronutrients including iron for bacterial disease management was hardly discovered in tomato plants.
In this study, the effect of exogenous iron (FeCl2) on in vitro growth of R. pseudosolanacearum and on bacterial wilt progression in detached tomato leaves were investigated.
R. pseudosolanacearum strain GMI1000 (formerly known as R. solanacearum GMI1000), causing tomato bacterial wilt, was isolated from tomato plants in French Guiana in 1978 (Deberdt et al., 2014; Tano et al., 2021). The bacterial strain was cultured in 4 ml of casamino acid-peptone-glucose (CPG) liquid medium in a 15-ml conical tube at 30°C overnight (Hong et al., 2016). The bacterial culture was centrifuged, and the bacterial pellet was resuspended with sterile water to adjust 0.15 at 600 nm using a spectrophotometer for a bacterial suspension (108 cfu/ml) (Hong et al., 2013). Different concentrations (5, 10, 20, 50, 100, 200, and 500 µM) of FeCl2 were added to 4 ml of the CPG broth in 15-ml conical tubes. The bacteria (105 cfu/ml) were initially inoculated in the CPG broth supplemented with the FeCl2, and cultured at 30°C with shaking until the OD value was reached 0.25±0.02 in the mock-treated control culture. Relative bacterial growths by FeCl2 treatments were expressed as percentage (%) compared to the mock-treated control culture. Independent experiments were performed four times, and each experiment contained four biological replicates. In vitro bacterial population was investigated after the liquid culture of R. pseudosolanacearum GMI1000 (Rp-GMI1000) in the absence or presence of FeCl2 (Fig. 1A). Increasing concentrations of FeCl2 suppressed the in vitro bacterial growth in dose-dependent manners. Fifty µM of FeCl2 began to reduce the bacterial growth (ca. 85.7%) compared to the mock-treated cultures, and the higher FeCl2 concentrations 100 and 200 µM more inhibited the bacterial growth to ca. 80.5 and 71.3%, respectively. Iron-reduced mycelial growth of the phytopathogenic fungi was shown, indicating the in vitro antifungal activity of exogenous iron. The mycelium dry weight of F. oxysporum were significantly reduced by more than 200 µ g/ml of FeCl3 (Hashem, 1995). Mycelial growth of six fungal species infecting grapevine, Botrytis cinerea, Eutypa lata, Phaeomoniella chlamydospora, Phaeoacremonium aleophilum, Diplodia seriata, and Neofusicoccum parvum, decreased on the media supplemented with FeSO4 (5 mM) (Fleurat-Lessard et al., 2011). However, direct antibacterial activity of iron has rarely demonstrated against phytopathogenic bacteria including R. pseudosolanacearum. Treatment with 50-200 µM of FeCl2 concentrations may suppress R. pseudosolanacearum by direct contact with the bacteria in this study, and it can be applied in the hydroponic cultures for many crops vulnerable to bacterial wilt. Both elevated bacterial inoculum levels and mechanical injury were involved in the increased bacterial wilt of tomato plants (Singh et al., 2014). The efficiency of the antimicrobial activity of FeCl2 against the increasing bacterial population of R. pseudosolanacearum more than 105 cfu/ml should be evaluated in a further study because 50-200 µM of FeCl2 in our current study limited the only bacterial numbers initially inoculated with 105 cfu/ml as an inoculum density.
Fig. 1.
Effect of iron on bacterial growth of Ralstonia pseudosolanacearum strain GMI1000 (Rp-GMI1000) and plant growth of detached tomato leaves. (A) Iron dose-dependent suppression of the in vitro bacterial growth. The bacterial suspension was initially prepared to the concentration of 105 cfu/ml in the liquid media supplemented with increasing doses (0, 2, 5, 10, 20, 50, 100, and 200 µM) of FeCl2 and shaking cultured at 30°C. The bacterial number was indirectly measured using a spectrophotometer with an optical density at 600 nm. Relative bacterial growth in the FeCl2-treated cultures was expressed as a percentage (%) compared to that in mock-treated cultures. (B) Phenotypes of the detached tomato leaves treated with different concentrations of FeCl2 (0, 2, 5, 10, 20, 50, 100, and 200 µM). Photos were taken at 3 and 5 days after petiole dipping in the different concentrations of FeCl2. (C) Relative fresh weight (%) of detached tomato leaves treated with different concentrations of FeCl2 after the petiole dipping at 3 and 5 days. Error bars represent standard errors of means of four independent experimental replications. Each experiment contained four biological replicates. Means with the same letter above the bars are not significantly different at the 5% level by the least significant difference tests.

Tomato seeds (cv. Cupirang) were sown in pots containing commercial soil mixtures and grown in a walk-in plant growth room under environmental conditions previously described in our study (Hong et al., 2018b). Third true leaves in the five-week-old plants were detached and used for chemical treatments and bacterial inoculation (Jo et al., 2020). We investigated phenotypic changes of the detached tomato leaves by dipping the petioles in the different concentrations of FeCl2 solutions because the excess iron in the ambient environment can lead to toxicity in diverse plant species (Ahammed et al., 2020; Li et al., 2016; Turhadi et al., 2019). Different concentrations (5, 10, 20, 50, 100, and 200 µM) of FeCl2 were prepared in distilled water. Distilled water was used as a mock treatment. The petiole of the detached tomato leaves was dipped in 10 ml of each solution in 50-ml conical tube. Fresh weight (FW) of the detached tomato leaves was measured before the petiole dipping in the FeCl2 solutions and then further measured at 3 and 5 days after the petiole dipping. Relative FW (%) of the FeCl2-treated leaves was expressed as a percentage compared to untreated leaves before the petiole dipping. Independent experiments were performed four times, and each experiment contained four biological replicates. Effects of FeCl2 applications on FW of the detached tomato leaves were investigated in the absence of the bacterial population (Fig. 1B, C). The distinct phenotypic change was not observed in the detached tomato leaves treated with increasing concentrations (2-200 µM) of FeCl2 at 3 and 5 days after petiole dipping, as shown in Fig. 1B. FW were investigated in the detached leaves at 3 and 5 d after petiole dipping (Fig. 1C). FW of the detached leaves were not different by the different concentrations of FeCl2 treatment at 3 and 5 days. These indicate that 2-200 µM of FeCl2 directly transported into the detached leaves through petioles did not cause a phytotoxic effect. Metal stresses to plants can depend on plant organs and growth stages (John et al., 2009; Souri et al., 2019). Therefore, we cannot exclude the possibility that treatment with FeCl2 (2-200 µM) in soil or hydroponic cultures can lead to damage in tomato roots. The FeCl2 concentrations can be re-adjusted to apply to the soil or hydroponic cultures of different growth stages of tomato plants.
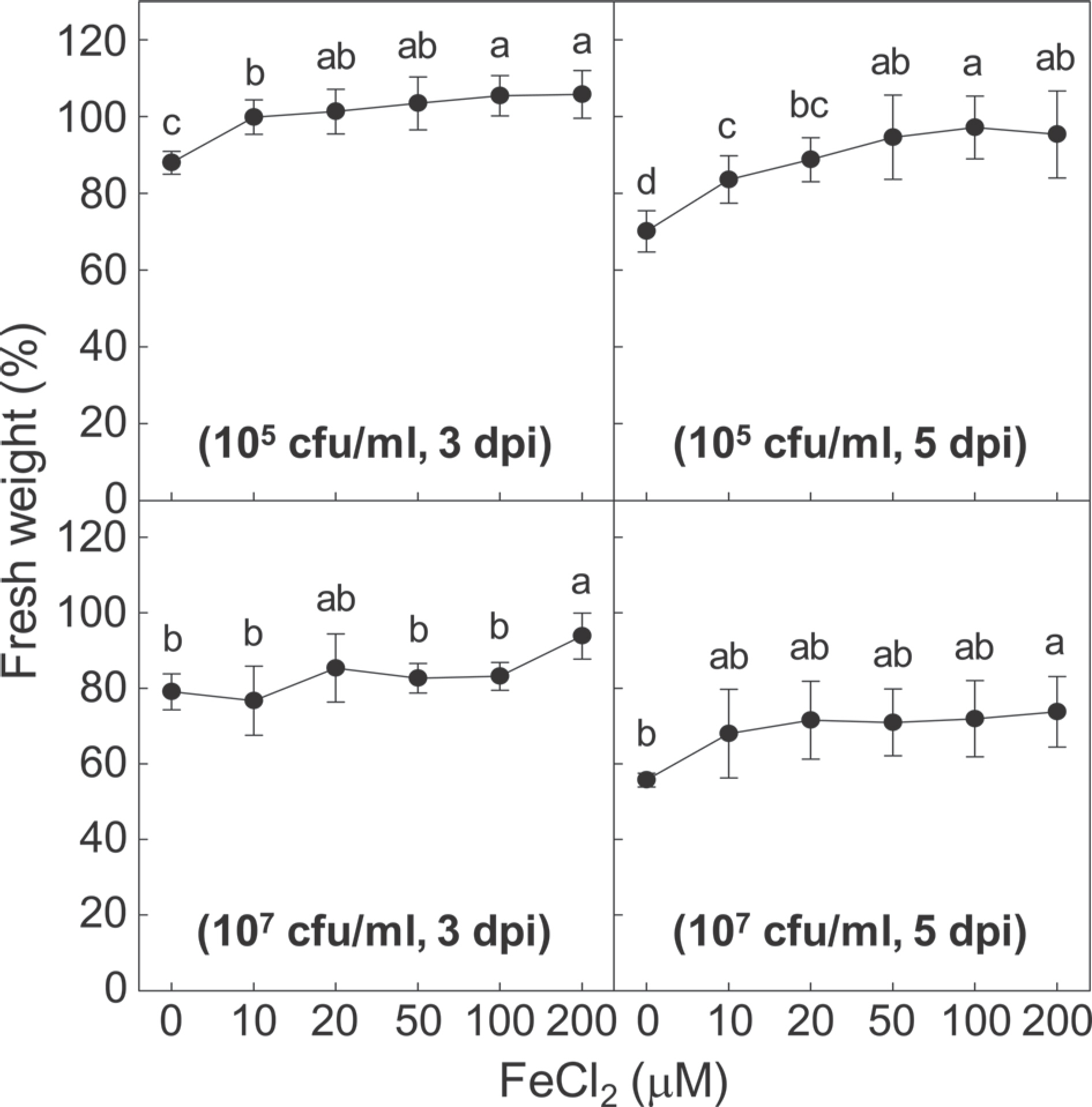
Petioles of the detached tomato leaves were placed in the 10-ml solutions of different FeCl2 concentrations and inoculated by bacterial suspension of R. pseudosolanacearum to make 105 and 107 cfu/ml as inoculum densities as described in our previous studies (Hong et al., 2013, 2018a). The bacterial wilt was evaluated by measuring the FW of the detached tomato leaves at 3 and 5 days after the bacterial inoculation. The FW of the detached tomato leaves at 3 and 5 dpi was compared to that of the leaves prior to the petiole dipping. Wilting symptoms of the detached leaves inoculated by the two inoculum densities (105 and 107 cfu/ml) of Rp-GMI1000 with or without FeCl2 treatments were demonstrated as relative fresh weights (%) (Fig. 2). Independent experiments were performed three times, and each experiment contained four biological replicates. Treatment with FeCl2 ameliorated the bacterial wilt of the detached leaves, and the reduced bacterial wilt was FeCl2 concentration-and the bacterium inoculum density-dependent. By the low inoculum density (105 cfu/ml), the reduced bacterial wilt was found in the leaves treated with more than 2 µM of FeCl2 at 3 dpi. The bacterial wilt suppression was more evident in the leaves treated with 100 and 200 µM of FeCl2 compared to the mock-treated control at 3 dpi. The detached leaves in the mock-treated control were more wilted at 5 dpi compared to 3 dpi. Increasing FeCl2 also reduced bacterial wilt at 5 dpi. The bacterial wilt was also suppressed in the leaves inoculated by the high inoculum density (107 cfu/ml) treated with different concentrations of FeCl2. In vitro antibacterial activity was shown by more than 50 µM of FeCl2 in this study, but the disease reduction in the detached leaves inoculated by Rp-GMI1000 (105 cfu/ml) could be achieved by 10-20 µM of FeCl2. It suggests that suppressed bacterial wilt of the detached tomato leaves treated with FeCl2 can be mediated by enhanced plant immunity together with direct antibacterial activity. Iron homeostasis has been closely associated with activating defences in several plants under pathogen threats, such as wheat leaves inoculated with Blumeria graminis f. sp. tritici (Aznar et al., 2015; Liu et al., 2007). Interestingly, iron homeostasis was important to regulate production of reactive oxygen species (ROS) in plant root development and leaf cell death (Reyt et al., 2015; Tewari et al., 2013). In our study, accumulation of ROS and lipid peroxidation were found in the petiole tissues of leaves inoculated by Rp-GMI1000 (Hong et al., 2013). Tomato leaf tissues directly syringe-infiltrated with R. solanacearum showed increased hydrogen peroxide generation in the bacterial dose-dependent manner (Flores-Cruz and Allen, 2009). Recently, hydrogen peroxide production and lipid peroxidation in the R. solanacearum-inoculated tomato leaves were significantly ameliorated during the silica nanoparticles-mediated disease control of tomato bacterial wilt (Wang et al., 2022). But hydrogen peroxide production in tomato leaves during the early phase of bacterial wilt was faster in high boron-treated protected plants (Jiang et al., 2016). The involvement of exogenous FeCl2 in redox regulation remains elucidated for understanding iron-mediated management of tomato bacterial wilt disease.
Fig. 2.
Effects of iron treatment on bacterial wilt disease of detached tomato leaves. Relative fresh weight of detached tomato leaves inoculated with two different Ralstonia pseudosolanacearum doses (105 and 107 cfu/ml) in the absence or presence of FeCl2 (10, 20, 50, 100, and 200 µM) was measured at 3 and 5 days post-inoculation (dpi). Error bars represent standard errors of means of three independent experimental replications. Each experiment contained four biological replicates. Means with the same letter above the bars are not significantly different at the 5% level by the least significant difference tests.
In conclusion, FeCl2 of more than 50 µM showed antibacterial activity against R. pseudosolanacearum. No adverse effect was found in the detached tomato leaves treated with FeCl2. The bacterial wilt of the detached tomato leaves was ameliorated by the FeCl2 application, and disease control efficacy was dependent on the FeCl2 concentration and bacterial inoculum density. Introducing microelement iron in tomato production fields can provide protection for tomato plants against bacterial wilt. Efficient FeCl2 application methods should be further evaluated in different growth stages of tomato plants in greenhouses for stable fruit production.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






