


서 론
근채류로서, 2018년을 기준으로 재배면적과 생산량은 2,154 ha와 73,143톤으로 보고되어 있다(Korea Statistical Information Service, 2020). 이런 당근을 재배하는 포장에서 생산성을 위협하는 여러 가지 병해 중에서 Alternaria dauci가 일으키는 당근 검은잎마름병은 전 세계적으로 큰 피해를 입히는 병이다(Netzer와 Kenneth, 1969; Strandberg, 1993; Vintal 등, 1999). 검은잎마름병이 발생하게 되면, 당근에서 광합성이 감소하게 되어, 품질 저하와 수확량 감소로 이어질 뿐만 아니라, 대규모 재배지에서 기계 수확할 때에는 지상부가 쉽게 잘려져 나가 수확량의 감소는 물론 노동의 곤란으로 이어지기도 한다(Ben-Noon 등, 2001; Dugdale 등, 2000; Strandberg, 1983). 국내에서도 2003년과 2004년에 국내 고랭지 재배지에서 당근 검은잎마름병이 70% 이상 심하게 발생하였고, 최근에는 가을 작형뿐만 아니라 봄 하우스 및 터널 작형에서까지 발생한다(Kwon 등, 2007). 국내 당근의 최대 주산지인 제주 지역에서도 당근 검은잎마름병은 주의해야할 병으로 알려져 있다. 병원균인 A. dauci는 최적 생장 조건에서 당근을 감염한 후 6-10일 안에 2차 전염원을 생성하며, 상대 습도가 낮을 때에는 생성된 포자가 바람을 타고서 다른 기주로 쉽게 전반되기 때문에 피해를 가속화시킨다(Langenberg 등, 1977; Strandberg, 1977). 또한, A. dauci의 포자는 내건성이 강하여 10% 정도의 포자는 수분 없이도 12시간까지 생존 가능할 뿐만 아니라, 오랜 기간동안 종자, 토양 혹은 병든 잎의 잔재물에서 월동할 수 있기 때문에 방제가 어렵다(Park 등, 2011; Pryor 등, 2002). 당근 검은잎마름병 방제 방법으로 수확 후 포장에서 병든 잎 제거, 토양의 질소 함량조절과 종자 온탕처리 등과 같은 경종적 혹은 물리적인 방제 방법이 시도되고 있지만, 가장 확실하고 신속하며 용이하게 방제할 수 있는 방법 중 하나는 살균제를 이용하는 화학적 방제 방법이다(Ben-Noon 등, 2001; Bounds 등, 2006; Koch 등, 2010; Kwon 등, 2007; Lee 등, 2011; Westerveld 등, 2008). 그러나 국내에서 당근 검은잎마름병을 살균제를 사용하여 방제하는 연구는 미미한 실정이다. 작물보호제지침서에는 같은 병임에도 불구하고 당근 검은잎마름병과 먹잎마름병으로 약제를 구분하여 표기함으로써 정보획득에 어려움을 겪게 하기도 한다(Korean Crop Protection Association, 2019). 또한 국내에 등록된 살균제는 chlorothalonil, propineb 등 보호 살균제 5종과 demethylation inhibiting계 2종으로 매우 단순하다. 등록된 살균제가 보호살균제인 ‘카’군과 탈메틸화효소의 활성을 억제하는 ‘사1’군만 있기 때문에 포장 살균제 처리 체계를 확립하고자 할 때, 살균제 선택이 매우 제한적이다. 또한 등록되어 있는 살균제의 대부분이 보호살균제이기 때문에 예방적으로 처리하는 시기를 맞추지 못하거나 초기 전염원 밀도가 높아 발병에 대한 심한 선택압을 받게 될 경우 방제 효과가 크게 감소할 수도 있다. 살균제 저항성 관리 차원에서도 작용기작이 다른 다양한 살균제가 사용되어야 한다.
따라서 본 실험에서는 작용기작이 다른 다양한 살균제를 선발하여 당근검은잎마름병균에 대한 in vitro상에서의 다양한 작용 특성을 조사하여, 당근 검은잎마름병의 방제에 사용할 수 있도록 제안하고, 또 주요한 살균제 그룹에 대한 포장 병원균의 저항성 모니터링을 실시하여 기존 살균제에 대한 감수성 기준을 정하고자 하였다.
재료 및 방법
실험에 사용한 균주와 균주의 보관.
실험에 사용한 당근검은무늬병균 Alternaria dauci KACC42997은 농업미생물보관센터에서 분양받아 사용하였다. 분양받은 A. dauci KACC42997은 potato dextrose agar (PDA) 배지에 접종하여 25°C의 암조건에서 10일간 배양하였다. 병원균 균총의 선단부에서 균사조각을 떼어내어 PDA 사면 배지에 접종하고 25°C에서 7일간 배양한 다음, 4°C에 보관하면서 실험에 사용하였다. 또한 장기간 보관하기 위하여 300 μg/ml의 streptomycin을 첨가한 멸균증류수를 각각의 cryotube vial (직경, 12 mm; 높이, 48 mm; Nunc Co. Ltd., Quebec, Canada)에 1.6 ml씩 담은 후, 배양한 균주의 균총 선단부에서 떼어낸 직경 3 mm의 균사조각을 떼어내어 cryo-tube vial에 각각 15조각을 넣고 상온에서 보관하였다.
살균제의 균사생장 억제 효과 조사.
여러 그룹의 살균제의 효과를 조사하기 위하여 보호살균제(‘카’군) 5종, 에르고스테롤 생합성 저해 살균제 중에서 ‘사1’군 6종과 ‘사3’군 1종, 세포호흡을 저해하는 ‘다2’군 살균제 5종과 ‘다3’군 살균제 4종 그리고 ‘다5’군 살균제 1종, 미세소관 형성 억제 살균제(‘나1’군) 3종과 ‘나1’군과 ‘나2’군의 혼합제 1종, 삼투압 신호 전달 저해 살균제(‘마2’와 ‘마3’군) 3종, 키틴 생합성 저해 살균제(‘아4’군) 2종, 병원균 세포막 합성 저해 살균제(‘바’군) 1종 등 총 32종의 살균제를 선발하여 당근검은잎마름병균 A. dauci KACC42997에 대한 균사생장 억제효과를 한천희석법으로 조사하였다. 세균 오염 방지를 위해 PDA배지에는 300 μg/ml의 streptomycin을 첨가하였으며, 배지에 첨가한 살균제의 최종 농도가 100, 10, 1, 0.1 μg/ml이 되도록 조절하였다. 한천희석법으로 살균제의 균사생장 억제효과를 조사하기 위해서 병원균을 25oC의 V-8 juice 배지(V-8 juice, 200 ml; CaCO3, 3 g; agar, 15 g; 증류수, 1 l)에서 10일간 배양한 후, 균총의 선단부에서 직경 3 mm의 균사조각을 떼어 정해진 농도 별로 살균제가 첨가된 PDA 배지에 접종하였다. 동일한 조건에서 10일간 배양 후, 배지의 균총의 직경을 조사하여 약제를 첨가하지 않은 무처리 PDA 배지의 균총 직경과 비교함으로써, 균사생장 억제율(%)을 다음의 식으로 계산하였다.
살균제의 포자 발아 및 발아관 신장 억제 효과.
실험에 사용한 32종의 살균제 중에서 보호 살균제 4종과 치료 살균제 8종을 선발하여 병원균의 발아 억제 효과를 실험하였다(Table 1).
A. dauci KACC42997의 포자 형성을 위해 병원균을 암조건의 25°C V-8 juice 배지에서 7일간 배양한 후, 멸균한 슬라이드글라스로 공중균사를 긁어 제거하였다. 공중균사를 제거한 병원균은 12시간 간격으로 근자외선을 조사하며 통기 상태의 20°C에서 7일간 배양하며 포자형성을 유도하였다. 형성된 포자는 150 μg/ml의 Tween 20 용액을 부어 수확한 후, 멸균된 거즈에 여과하여 균사조각을 제거한 포자 현탁액을 얻었다. 포자 현탁액은 3,000 rpm으로 20분간 2회 원심분리하여 세척 후, 다시 멸균수에 현탁하여 포자의 최종 밀도가 5×104 개/ml가 되도록 조절하였다. 포자 발아와 발아관 신장은 살균제를 첨가한 PDA 배지 위에 멸균된 셀로판막을 올려놓고 실시하였다. PDA 배지에 300 μg/ml의 streptomycin과 선발한 각 살균제의 최종농도가 100, 10, 1 μg/ml이 되도록 첨가하였다. 살균제를 처리하지 않은 무처리 PDA 배지와 처리한 PDA 배지 위에 동일한 크기(1.5 cm×1.5 cm)로 잘라 멸균한 셀로판막을 올려놓은 후, 한 장의 셀로판막 위에 준비한 포자 현탁액을 150 μl씩 점적하였다. 병원균을 접종한 PDA 배지는 포화습도가 유지되는 플라스틱상자에 넣어 25°C 암조건에서 배양하였으며, 모든 처리는 3반복으로 실시하였다. 살균제의 포자발아 억제율은 1반복당 100개의 포자를 조사하여 발아율을 구한 후 아래의 식으로 계산하였다. 이 때 포자의 발아 여부는 발아관이 포자의 단경보다 길게 발아한 경우만 발아한 것으로 판단하였다. 또한 각 처리구의 반복당 발아된 50개의 포자에서 발아관의 길이를 조사하였으며, 다음 식으로 발아관 신장 억제율(%)을 조사하였다. 발아관 길이는 포자에서 나온 발아관 중 단경보다 긴 발아관의 길이를 모두 재어 평균값을 구하였다.
살균제의 포자 형성 억제 효과.
A. dauci KACC42997을 직경 9 cm의 페트리접시에 부어 준비한 V-8 juice 배지에 접종하여 25oC의 암조건에서 7일간 배양한 후, 균총의 직경이 75 mm 이상이 된 배지만을 선발하여 멸균된 슬라이드글라스로 공중 균사를 제거하였다. 선발한 살균제 농도를 100 μg/ml로 조절한 다음, 살균제 용액 150 μl를 균총 전면에 고르게 점적하여 처리하였다. 살균제를 처리한 배지는 20°C의 통기 조건에서 12시간 간격으로 근자외선을 조사하며 7일간 배양하며 포자 형성을 유도하였다. 형성된 포자는 10 ml 멸균수를 부어 수확한 후 2겹의 멸균한 거즈에 여과하여 균사체 등을 제거하였다. 수확한 포자현탁액의 포자 밀도는 혈구계산기를 이용하여 조사하였으며, 무처리구와 비교하여 다음과 같은 식으로 포자 형성 억제율을 계산하였다.
결과 및 고찰
병원균 균사생장에 대한 살균제의 억제 효과.
실험에 사용한 5종의 보호살균제(‘카’군) 중에서 iminoctadine tris-albesi-late를 제외한 나머지 4종의 EC50값은 모두 1.5 μg/ml 이상으로 효과가 저조하였다(Table 2).
이러한 결과는 Stepanović 등(2015)의 실험 결과와 동일하였다. 토마토에서 겹둥근무늬병을 일으키는 A. solani에 대해서 보호살균제인 chlorothalonil과 치료살균제인 defeno-conazole의 균사생장 억제효과를 한천희석법을 사용하여 조사한 결과, chlorothalonil의 EC50값은 2.99-4.54 μg/ml이었고, difenoconazole의 EC50값은 0.018-0.037 μg/ml로, 보호살균제의 병원균에 대한 균사생장 억제효과가 ‘사1’군에 속하는 살균제보다 저조하였다. 하지만 동일한 ‘카’군에 속하는 iminoctadine tris-albesilate의 EC50값은 0.185 μg/ml로 우수하였는데, 저장 감귤의 부패에 관여하는 Penicillium italicum과 P. digitatum에 대한 균사생장 억제효과를 조사한 Hyun 등(2001)의 결과에서도 P. italicum과 P. digitatum에 대한 iminoctadine tris-albesilate의 EC50값은 각각 0.01-0.02 μg/ml와 0.01 μg/ml로, 균사생장 억제효과가 우수하였다. 하지만 고추흰비단병균에 대한 iminoctadine tris-albesilate의 균사생장 억제효과는 EC50값이 100 μg/ml 이상으로 매우 저조하였다(Lee 등, 2017). 이처럼 살균제의 효과는 적용하는 병원균에 따라서 효과가 다르게 나타나기 때문에, 특정 살균제의 적용 병해 확대를 위해서는 살균제의 효과 검정을 정확하게 수행한 후 결정해야 할 것으로 생각한다.
세포 호흡을 저해하는 ‘다’군 살균제는 미토콘드리아 전자전달계의 복합체 II를 구성하는 숙신산 탈수소 효소 기능 저해제(‘다2’군) 5종, 복합체 III을 구성하는 cytochrome bc1 효소의 기능 저해제(‘다3’군) 4종, 그리고 산화적 인산화 반응 저해제(‘다5’군) 1종의 살균제를 선발하여 균사생장 억제효과를 조사하였다. ‘다2’군에 속하는 살균제 중에서 isopyrazam, fluxapyroxad, boscalid, fluopyram의 EC50값은 0.579, 0.053, 0.155, 1.443 μg/ml이었으나, 동일한 작용기작을 갖는 fluto-lanil은 100 μg/ml 이상으로 효과가 저조하였다. Flutolanil은 Rhizoctonia solani의 숙신산 탈수소효소의 기능을 저해하여 균사생장을 억제하는 살균제로 알려져 있지만(Hirooka 등, 1990a, 1990b), 담자균류 이외의 자낭균이나 난균문에 속하는 병원균에 대한 효과는 매우 미미하며 숙신산 탈수소효소의 기능도 전혀 억제하지 않는다(Motoba 등, 1988). 본 연구결과에서도 자낭균문에 속하는 A. dauci에 대해서는 EC50값이 100 μg/ml 이상으로 균사생장 억제효과가 아주 미미하였다. ‘다3’군에 속하는 cytochrome bc1 효소 활성 억제 살균제 중에서 pyraclostrobin의 EC50값만이 1.174 μg/ml이었으며, 나머지는 모두 30 μg/ml 이상으로 균사생장 억제효과가저조하였다. ‘다3’군에 속하는 strobilurin계 살균제는 병원균의 포자발아 억제효과에 비하여 균사생장 억제효과가 떨어진다(Steinfeld 등, 2001). 따라서 한천희석법을 이용한 살균제 효과 검정에서는 포자 발아를 억제하는 살균제의 효과는 저조하게 나올 수 있기 때문에, 살균제의 특성을 정확하게 검정할 수 있는 방법으로 살균제의 실내 검정을 실시하여야 한다. 미토콘드리아 내막에서 H+ 이온의 이동을 억제하는 ‘다5’군에 속하는 fluazinam의 균사생장 억제효과에 대한 EC50값은 0.083 μg/ml로 우수하였다. 동일하게 호흡을 저해하는 살균제라고 하더라도 병원균의 종류와 미토콘드리아의 전자전달과정에서 활성을 억제하는 작용점 효소가 달라짐에 따라서 균사 생장을 억제하는 효과는 크게 달라졌다. ‘다2’군의 살균제에서 보는 것과 같이 동일한 효소를 작용점으로 가지는 경우에도 살균제의 화학 구조에 따라서 효과는 달라졌다. 효과가 우수하였던 isopyrazam과 fluxapyroxad는 pyrazole-4-carboxamide계에 속하며, boscalid는 pyridine-carboxamide계에 속하지만, 효과가 감소한 fluopyram은 pyridinyl-ethyl-benzamide계에 속하였고, flutolanil은 phenyl-benzamide계의 살균제였다.
에르고스테롤 생합성을 억제하는 ‘사’군 살균제에서는 탈메틸효소 활성저해 살균제(‘사1’군) 6종과 케토 환원 효소의 활성을 저해하는 살균제(‘사3’군)인 fenhexamid 1종을 선발하여 실험하였다(Table 2). 탈메틸 효소의 활성을 억제하여 에르고스테롤 생합성을 억제하는 살균제는 metconazole의 EC50값이 1.055 μg/ml이었을 뿐, 나머지 5종 살균제의 EC50값은 모두 1.0 μg/ml 이하로 A. dauci의 균사생장을 크게 억제하였다. 하지만 케토 환원 효소의 활성을 저해하는 fenhexamid의 EC50값은 2.631 μg/ml로, ‘사1’군에 속하는 살균제의 균사생장 억제효과보다 저조하였다. ‘사1’군의 살균제 중에서 효과가가장 우수하였던 살균제는 difenoconazole로서 EC50값이0.002 μg/ml이었다.
신호전달을 저해하는 ‘마’군에 속하는 살균제 중에서 ‘마2’군이며 phenylpyrrole계인 fludioxonil의 EC50값은 0.017 μg/ml이었으며, ‘마3’군인 dicarboximide계인 iprodione과 procymi-done의 EC50값은 각각 0.414와 1.926 μg/ml이었다. 미세소관 형성을 억제하는 4종의 benzimidazole계 살균제와, 키틴 합성 저해제인 polyoxin, 지질 생합성 저해제인 isoprothiolane의 EC50값과 minimum concentration (MIC) 값은 모두 100 μg/ml 이상으로, 당근검은잎마름병균인 A. dauci KACC42997에 대한 균사생장 억제효과가 매우 미미하였다. Table 2에서 보면 iminocta-dine tris-albesilate처럼 EC50값이 낮은데도 불구하고 MIC값은 >100 μg/ml인데, EC50값이 1.701 μg/ml로 iminoctadine tris-albesilate보다 큰 propineb의 MIC값은 100 μg/ml이었다. 이런결과는 살균제에 따라서 충분히 나타날 수 있는 결과이기 때문에, 균사생장 억제효과를 조사하여 살균제의 효과를 비교하고자할 때에는 EC50값과 MIC값을 동시에 조사하여 비교하는 것이타당하다.
살균제의 포자 발아 및 발아관 신장 억제 효과.
A. dauci KACC42997을 PDA 배지 위에 올려놓은 셀로판막 위에 접종하고 25oC에서 배양하면, 1.5시간 배양한 후부터 병원균 포자의 각각의 세포에서 발아관이 생성되며 발아하기 시작하였다(Fig. 1).
배양 시간별로 포자 발아 상태를 확인하였을 때, 3시간 후에 이미 97.2%의 포자가 발아하였으며, 6시간 후에는 100.0% 포자가 발아하고, 발아관이 분지하며 자라기 시작하였다. 병원균의 포자를 9시간 동안 배양한 후에는 다세포 포자에서 생성된 여러 개의 발아관이 서로 얽혀 자라기 때문에 발아관의 길이를 조사가 용이하지 못하였다. 따라서 보호 살균제 4종과 치료 살균제 8종의 포자 발아 억제율과 발아관 신장 억제율은 살균제 배지에 올려놓은 셀로판막 위에 포자 현탁액을 접종하고 6시간 동안 배양한 후에 포자 발아율과 신장한 발아관의 길이를 조사하여 계산하였다. 살균제 무처리구의 포자는 100%가 발아하였으며, 발아관 길이는 평균 226.01 μm이었다. ‘카’군 살균제에 속하는 chlorothalonil, dithianon, folpet, iminoctadin tris-albesilate 중에서 chlorothalonil, dithianon, folpet은 병원균에 대한 포자발아 억제효과가 우수한 반면에 균사생장 억제효과는 낮았으나, iminoctadin tris-albesilate는 반대로 포자 발아 억제효과는 낮았지만 균사생장 억제효과는 우수하였다(Table 3). Cytochrome bc1 복합체의 활성을 저해하여 호흡을 저해하는 다3군에 속하는 pyraclostrobin은 병원균의 균사생장뿐만 아니라 포자발아에 대한 억제효과도 우수하였다. 하지만 pyraclostrobin의 균사생장 억제효과와 포자발아 억제효과를 EC50값으로 비교하여 보면, 균사생장에 대한 EC50값은 1.763 μg/ml이고 포자발아에 대한 EC50값은 0.001 μg/ml 이하로, 균사생장에 대한 억제효과보다는 포자발아에 대한 억제효과가 훨씬 우수함을 알 수 있었다. 세포 호흡에서 산화적 인산화 반응을 저해하는 다5군에 속하는 fluazinam은 병원균의 균사생장뿐만 아니라 포자발아에 대한 억제효과도 우수하였는데, 1.0 μg/ml의 농도에서도 포자발아와 균사생장을 각각 97.3%와 82.7% 억제하였다. 발아관 신장 억제효과는 균총의 길이를 측정하여 조사한 균사생장 억제효과와 높은 상관관계를 보이면서, 균사생장 억제효과가 우수하였던 살균제의 발아관 신장 억제효과도 우수하였다(Table 3).
병원균의 포자 형성 억제 효과.
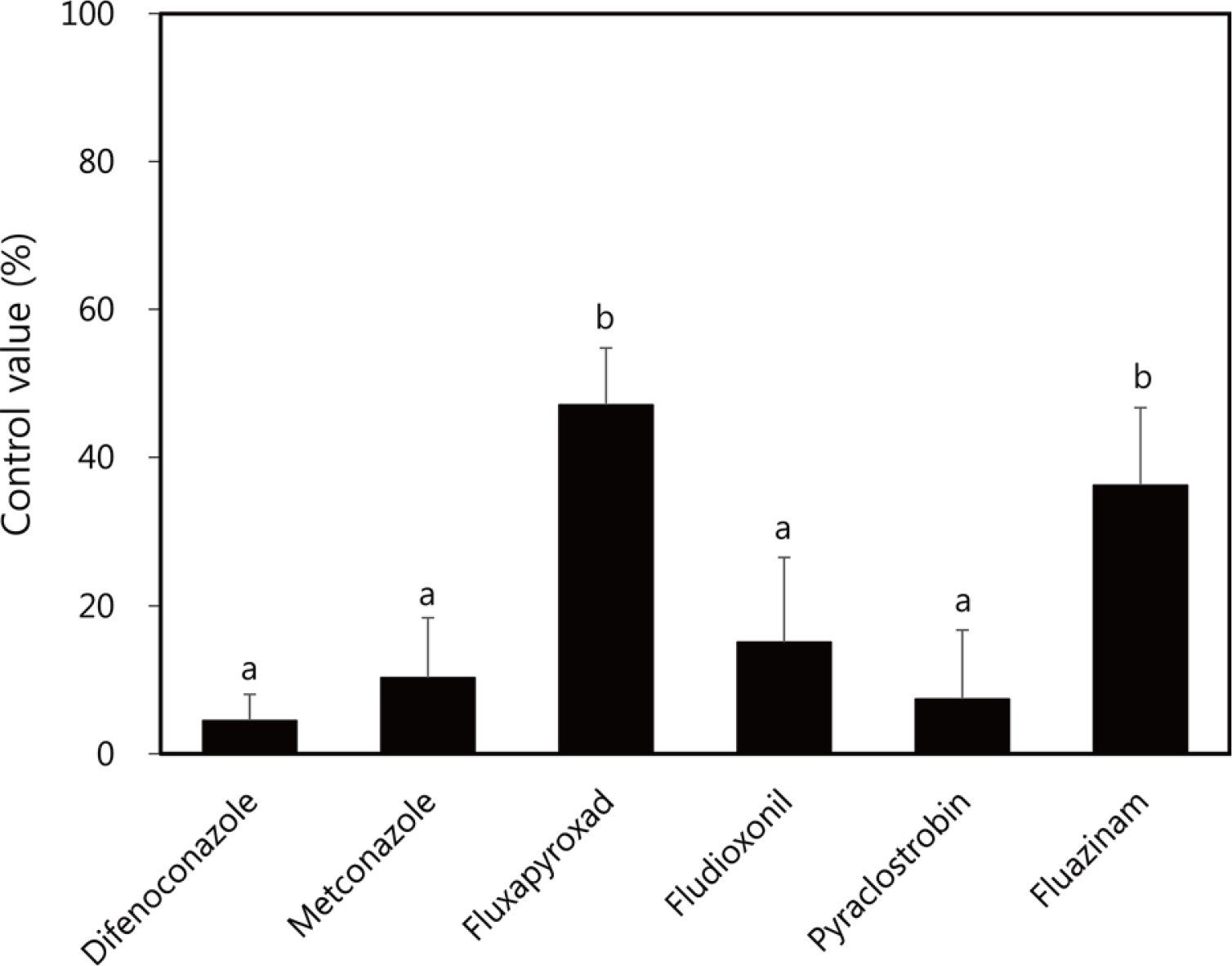
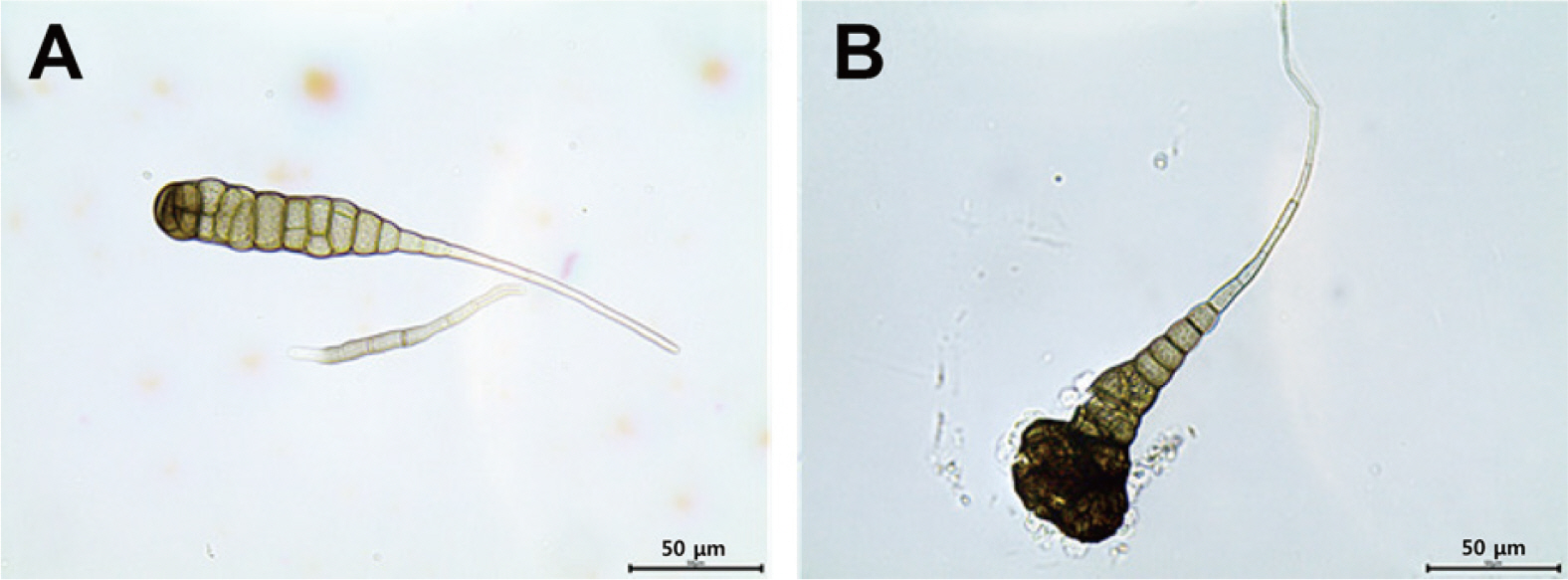
6종의 살균제 중에서 100 μg/ml의 fluxapyroxad를 처리한 배지에서 생성된 포자의 수가 무처리구에서 생성된 포자 수보다 47.1% 억제되었으며, fluazi-nam 처리는 36.2%를 억제되었다(Fig. 2). 그 외 살균제는 무처리구의 포자 형성량과 거의 차이가 없었다. 다만 fludioxonil 처리구의 경우 포자 형성량에는 큰 차이가 없었으나, 포자의 형태가 일부 변형되어 있었다(Fig. 3). 이처럼 포자가 변형되는 원인은 fludioxonil의 작용기작과 관련이 있을 것으로 추정한다.
Fludioxonil은 병원균의 삼투압에 대한 신호전달을 저해하는
살균제로 알려져 있는데, 이런 작용기작이 새롭게 생성되는 A. dauci의 포자를 형태적으로 변형시켰을 것으로 추정한다. 따라서 이런 추론을 증명하기 위한 생화학적인 실험이 뒷받침되어야 할 것으로 본다(Fig. 3).
각 지역 병원균 집단의 살균제에 대한 감수성 차이.
에르고스테롤 생합성 저해 살균제에 대한 병원균 집단의 모니터링 결과, prochrolaz의 EC50값은 0.158 μg/ml (0.002-0.476 μg/ml)로, 실험한 살균제 중에서 병원균의 균사생장을 가장 효과적으로 억제하였다. 또한 당근 검은잎마름병 방제를 위해서 등록되어 있는 살균제인 difenoconazole과 metconazole도 0.219 μ g/ml (0.018-0.946 μg/ml), 0.350 μg/ml (0.042-1.683 μg/ml)로 병원균의 균사생장 억제효과가 우수하였다. 그러나 에르고스테롤 생합성 저해 과정 중 C-4 위치의 탈메틸화 과정에서 3-keto reductase의 활성을 저해하는 fenhexamid의 경우, ‘사1’ 군에 속하는 다른 살균제에 비해서 균사생장 억제효과가 낮았다. Table 4에서 보는 것과 같이 각 지역 별 병원균 집단의 살균제 반응에는 차이가 있었다. 구미 지역 병원균 집단은 difeno-conazole과 prochloraz에 대한 Rf(저항성 요인)값이 다른 지역보다 크게 나타나, 병원균 집단 내에서 살균제에 대한 감수성정도가 타 지역보다 다양하다는 사실을 알 수 있었다. Tebuco-nazole에 대해서도 구미 지역 집단은 제주 지역 집단과 대등한 평균 EC50값을 보이면서도 Rf값이 큰 것을 보면 집단 내 각 균주 간의 감수성 정도가 다양한 것을 알 수 있다. 평창 지역의 집단은 metconazole과 tebuconazole에 대해서 평균 EC50값뿐만 아니라 Rf값마저 높게 나타나, 지역 병원균 집단에 속하는 A. dauci 균주의 살균제에 대한 감수성이 전체적으로 저하되었을 뿐만 아니라 집단 내의 다양성 역시 높음을 보여주었다. 하지만 fenhexamid에 대한 각 지역 병원균 집단의 EC50값과 Rf값은 비슷하였다. 조사한 세 지역 중에서 제주 지역 병원균 집단이 살균제에 대한 감수성 정도의 다양성이 가장 낮았다. Rf(저항성 요인)값이 크다는 의미는 지역 내 병원균 집단의 감수성 정도에 대한 다양성이 높다는 것을 뜻하는데, 집단 내에서 각 균주의 감수성 정도가 다르기 때문에 그만큼 저항성 발현이 용이한 상태임을 말하고 있다.
실험에 사용한 모든 A. dauci 균주는 숙신산탈수소효소를 저해하는 isopyrazam과 fluxapyroxad에 대한 평균 EC50값이 2.350과 fluxapyroxad 1.644 μg/ml이었다. 그러나 숙신산탈수소효소의 활성을 저해하면서도 isopyrazam이나 fluxapyroxad와는 화학구조가 다른 boscalid와 complex III에 작용하여 ATP 합성을 저해하는 pyraclostrobin에 대한 EC50값은 높았다(Table 5). 검은잎마름병균의 지역별 감수성 차이를 비교했을 때, 구미와 평창보다 제주의 Rf값이 적게 나타나, 제주 지역 병원균 집단의 살균제 감수성 정도는 다른 지역보다 다양성이 낮다는 것을 알 수 있었다. 이런 결과는 제주 지역이 다른 지역보다살균제 사용이 많지 않았을 가능성을 보여준다. A. dauci에대한 각 살균제 반응의 상관관계를 조사해 보면 isopyrazam과fluxapyroxad, isopyrazam과 boscalid에서 0.01 수준으로 유의한 상관관계가 나타났다(Table 6). 하지만 pyraclostrobin은 나머지 3개 살균제와는 상관관계가 없었다.
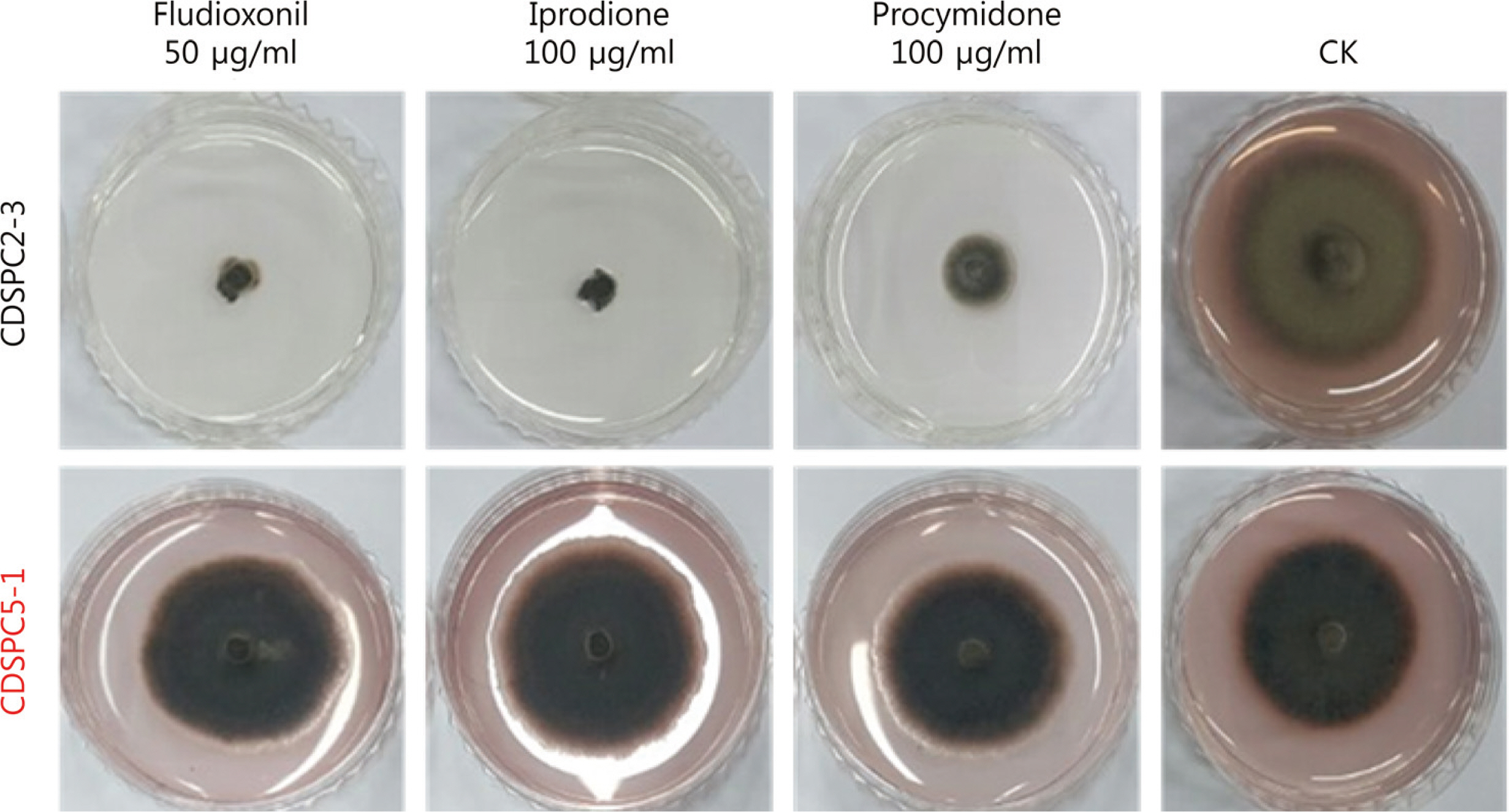
Phenylpyrrole계에 속하는 fludioxonil(‘마2’군)에 대한 A. dauci 반응을 조사하였을 때, 평창 포장에서 2개의 저항성 균주가 발견되었다(Fig. 4). 이 두 저항성 균주는 dicarboximide계(‘마3’군)에 속하는 iprodione과 procymidone에 대해서도 높은 저항성을 보였다. Fig. 5에서 보는 것과 같이 감수성인 CD-SPC2-3 균주는 50 μg/ml의 fludioxonil과, 100 μg/ml의 iprodi-one과 procymidone이 첨가된 PDA 배지에서 생장이 크게 억제되었지만, 저항성인 CDSPC5-1 균주는 살균제를 첨가한 배지에서의 생장이 살균제를 첨가하지 않은 PDA 배지에서 생장과 차이가 없었다.
이결과는 A.dauci가 phenylpyrrole계의 fludioxonil과 dicar-boximide계에 속하는 iprodione과 procymidone 사이에 교차저항성이 존재함을 보여 주고 있다. 저항성 균주인 A. dauci CD-SPC5-1과 CDSPC5-2를 제외하고 지역 별로 각 균주의 EC50값을 구하여 비교한 Rf값을 보면, 다른 기작을 지닌 살균제의 경우와 동일하게 제주 지역 병원균 집단의 Rf값이 여전히 낮았다. 제주 지역에서는 당근을 유기농으로 재배하는 곳이 많기 때문에 살균제의 사용이 적어서 타 지역에 비하여 적었기 때문이라고 생각한다.
요 약
당근검은잎마름병균인 Alternaria dauci KACC42997에 대해서 32개 살균제를 선발하여 균사생장 억제효과를 조사한 결과, ‘다2‘군, ’다5‘군, ’사1‘군, ’마2‘군, ’마3‘군에 속하는 살균제의 균사생장 억제효과가 우수하였다. Iminoctadine tris-albesilate를 제외한 ‘카’군에 속하는 보호살균제와 ’다3‘군에 속하는 pyra-clostrobin은 병원균의 포자발아를 억제하는 효과가 우수하였다. 균사생장 억제효과가 우수했던 숙신산탈수소효소 활성을 저해하는 ’다2‘군과 탈메틸효소 활성을 저해하는 ’사1‘군 살균제는 우수한 균사생장 억제효과를 보여주면서도, 포자발아 억제효과는 저조하였다. 하지만 ’다5‘군에 속하는 fluazinam은 균사생장 억제효과뿐만 아니라 포자발아 억제효과도 우수하였다. 특별히 ’다2‘군에 속하는 fluxapyroxad를 100 μg/ml 처리한 경우, 배지 상에서 포자형성을 47.1% 억제하였다. 구미, 평창, 제주 등에서 분리한 검은잎마름병균 집단의 ’사‘군, ’다‘군, ’마‘군 살균제에 대한 저항성 요인값을 비교하면, 제주 지역 병원균 집단의 저항성 요인값이 가장 낮았다. 평창 지역 균주 집단에서 ’ 마2‘군에 속하는 fludioxonil 저항성 균주가 2개 발견되었으며, 그들은 모두 iprodione과 procymidone에 대해서 교차저항성을 보였다.


 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






