



Introduction
Plantation forests are native and introduced tree species planted primarily for wood biomass, timber, wood, pulp, and environmental enhancement activities around the world. Eucalyptus, Acacia, and pine are the most commonly planted tree species worldwide (Tadesse et al., 2019). According to Food and Agriculture Organization (2020), the global, Afri-can, and Ethiopian total area of planted forests is estimated to be 7%, 67%, and 80%, respectively. Ethiopian plantation forests consist of introduced plant species of Eucalyptus, Cupressus, Casuarina, Juniperus, and Pinus (Mesfin and Wubalem, 2014). According to Dessie et al. (2019), more than 55 species of Eucalyptus have been used for plantation in Ethiopia. Ethiopia is one of the 10 countries with the largest area under Eucalyptus plantation (506,000 ha) in East Africa (Dessie and Erkossa, 2011; Lemenih and Kassa, 2014). Rapid growth, coppice availability, and ease of management allow Eucalyptus species to be grown on a large scale, from households, and small-scale private plantations, to large state forest enterprises in Ethiopia (Mesfin and Wubalem, 2014). Eucalyptus plants were introduced primarily to alleviate the shortage of wood biomass in Ethiopia, but are now widely used for construction, poles, firewood, charcoal, plywood, building materials, fence posts, rails, and alternative sources of income (Dessie et al., 2019; Food and Agriculture Organization, 2009).
According to the report of Slippers and Wingfield (2007), the establishment of plantations with exotic species pro-vides the opportunity for the growth, development, and spread of pathogens around the world. According to Food and Agriculture Organization (2020), the average area of forest affected by the disease from 2002 to 2017 was estimated at 4.76 million ha in 37% of the world. The report shows that the impact of diseases on forest production is increasing significantly in the world. Research findings revealed that Eucalyptus plant species in different agroecological areas around the world had been damaged by various fungal and bacterial plant pathogens (Chungu et al., 2010). Several studies have shown that fungi belonging to the Aplosporellaceae, Mycosphaerellaceae, and Teratosphaeriaceae families are the cause of diseases that seriously affect the productivity of Eucalyptus plantations around the world (Gezahgne et al., 2003; Mohali et al., 2007; Old et al., 2003). According to Wingfield et al. (2008), the causal pathogens for Eucalyptus plant diseases originate either from native areas of host tree species or cross-infection from native plants of importing countries.
Disease reports on exotic tree plantations in Ethiopia by Darge (2017), Darge and Bogale (2017), Demissie et al. (2020), and Gezahgne et al. (2003, 2005, 2006), confirm, severe association, incidence, and distribution of leaf spot and stem canker diseases caused by Mycosphaerella, Botryosphaeria, Cylin-drocladium, Chrysoporthe, Teratosphaeria, and Phoma fungal species in Ethiopia. Most of these studies are conducted in a limited area of the country and do not involve molecular or phylogenetic studies for identification. Limited research and a knowledge gap on the diseases that affect the productivity of Eucalyptus plantations in Ethiopia are key initiatives for this study. The main objective of this study was to evaluate the diseases currently associated with Eucalyptus plants and to identify causal fungal pathogens to provide up-to-date infor-mation and contribute to the development of appropriate disease management options.
Materials and Methods
Study site description.
The study was conducted in the Eucalyptus growing areas of Bedelle (BE), Jima (JI), Abru (AB), Woreta (WO), Humbo (HU), Ana lamo (AN), and Masken (MA), districts in Oromia, Amhara and Southern Nations Nationalities and Peoples (SNNP) regions of Ethiopia. In the Amhara region, the average annual minimum and maximum tem-peratures range from15°C and 21°C, the average annual rainfall is 1,194 mm, and the altitude ranges from 500 to 4,620 meters above sea level (Bewket, 2009). The average annual minimum and maximum temperatures; and rainfall ranged from 18°C to 39°C and 450 to 820 mm, respectively, while the altitude ranged from 500 to 4,607 above sea level in the Oromia region (Bewket, 2009). In similar cases, the mean annual rainfall in the SNNP region ranges between 200 and 2,200 mm, and the mean annual temperature ranges from 15°C to 26°C while the altitude range in the region is 375 to 4,207 meters above sea level (Gebremichael et al., 2014). Details of the study areas are given in Fig. 1.
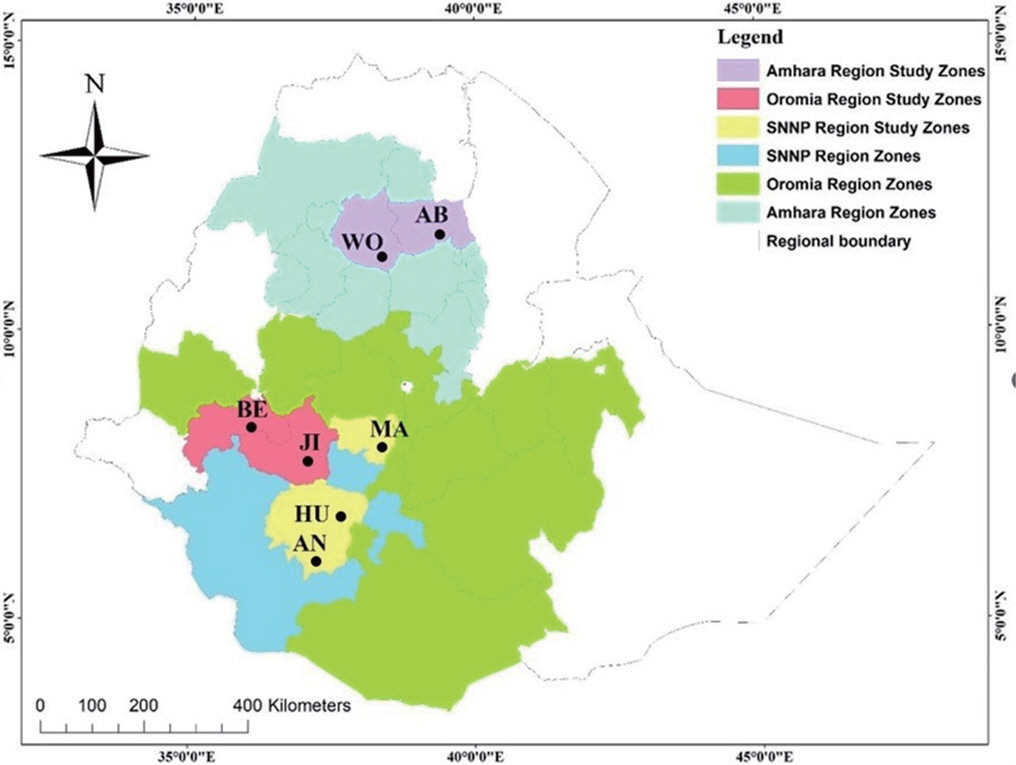
Survey and sampling from Eucalyptus plant fields.
The disease survey was conducted in the Eucalyptus plantation areas in Ethiopia during 2019/2020 and 2020/2021. The survey and sampling were carried out at nine locations in the regions. Two plantation sites located 3‒5 km apart, were randomly selected for disease assessment. Plantation sites were selected on the basis of agroecology, road access, and productivity. Five Eucalyptus trees were randomly selected from each plantation and examined for disease symptoms. Samples were collected from the Eucalyptus plant with symptoms of cracking of the stem and branches, cankers, and tip dieback as described by Li et al. (2018), Masood et al. (2010), and Pillay et al. (2013). The collected samples were placed in paper bags and transported to the laboratory for identification.
Disease assessment.
The incidence and severity of stem and branch canker disease were determined using the disease index described by El-Gali and El-Zahaf (2015) and Masood et al. (2010). The percentage of disease incidence was calculated using the following formula: Percentage of the canker disease incidence=(No. of diseased trees/Total No. of trees)×100.
Disease severity of stems and branch canker was determined using a scale of 0‒7, where 0=no sign of canker, 1= 1‒10% of canker over the entire observed area, 2=11‒20%, 3=21‒30%, 4=31‒40%, 5=41‒50%, 6=51‒60%, and 7=61% and above were considered as maximum canker severity. Disease severity was calculated using the formula:
Percentage of the canker diseases severity=(Σ(n×v)×100)/(N×S), where Σ=Sum, n=number of trees in each category, v= numerical value of the observed trees, N=total number of examined trees, and S=the maximum canker severity numerical value.
Isolation, characterization, and identification of fungi from diseased samples.
For the isolation and purification of fungal cultures, diseased sample segments were sterilized in 70% ethyl alcohol for 1 min and in 2% sodium hypochlorite for 2 to 4 min, washed three times with sterile distilled water, dried on sterile filter paper, aseptically transferred to malt extract agar (MEA) and kept at room temperature (25‒27°C) for 5 to 15 days (Ahmadpour et al., 2017). The hy-phal tip transfer method was used to subculture and obtain pure isolates. Alternatively stems and branches with clear cankers symptoms were sequentially surface sterilized in 70% ethyl alcohol for 1 min and in 2% sodium hypochlorite for 2 to 4 min, washed three times with sterile distilled water, dried on sterile filter paper and incubated in moist chambers to induce the development of a fungal fruiting body. After appearance and maturity of fruiting bodies monosporic cultures were generated by plating a spore taken from pycnidia suspended in sterile water on MEA. Pycnidia and conidia produced on stems and branch samples were mounted on microscope slides using distilled water and the structures of conidiomata, ascospores, and conidia were examined under the Olympus BX53 digital fluorescence microscope (Tokyo, Japan) as described by Chen et al. (2015).
Isolates were cultured and subcultured to obtain the desired purity for morphological studies. In cultures, characteristics such as colony diameter, color, texture, concentric zone, fruiting body shape, and conidia; shape, septation, pigmentation, and size were studied (Aveskamp et al., 2010; Noordeloos et al., 1953; Silva et al., 2020). All pure fungal isolates were classified according to their morphological characteristics and finally identified at the species level using internal transcribed spacer (ITS) rDNA sequence analysis.
DNA isolation, amplification.
DNA extraction was performed from 10-day-old cultures of fungal isolates, as described by Li et al. (2020). The harvested fungal mycelia were pulverized in liquid nitrogen, lysed with buffer, then dis-solved in phenol, chloroform, and isoamyl alcohol, and finally, the desired DNA was recovered by an ethanol precipitation method. The DNA base concentration was determined using a NanoDrop spectrophotometer (ND-1000, NanoDrop Technologies, Wilmington, DE, USA). To measure the quality of the obtained DNA, electrophoresis was performed on a 1% agarose gel (Raja et al., 2017). The ITS region of the DNA sample was amplified using the first internal transcribed (ITS1) and the second ITS region (ITS2) primers as described by White et al. (1990).
Sequencing and phylogenetic analyses.
The poly-merase chain reaction (PCR) products were cleaned using a High Pure PCR Product Purification Kit (Roche Molecular Biochemicals, Mannheim, Germany) and sequenced in both directions with the same primers as used for PCR. The ABI PRISM Big Dye Terminator Cycle Sequencing Ready Reaction Kit (Perkin-Elmer Applied Biosystems, Foster City, CA, USA) was used to perform the sequencing reactions and the sequences were run on an ABI PRISM 377/3100 Autose-quencer (Perkin-Elmer Applied Biosystems) as described by White et al. (1990) at the Netherlands Microbiology Research Centre (MRC) and the Center for Agriculture and Bioscience International (CABI), UK using forward and reverse primers. ITS sequences obtained were trimmed using BioEdit v. 7.0.9.0 (Hall, 1999), and alignments for each sequence were determined using the Basic Local Alignment Search Tool (BLAST) in the NCBI database (National Center for Biotechnology In-formation, National Library of Medicine, Bethesda, MD, USA) for identification. Consensus sequences were assembled in Molecular Evolution Genetic Analysis (MEGA) version 10 software (Kumar et al., 2018). Additional known sequences of genera Didymellaceae and outgroup were obtained from GenBank and the phylogenetic tree was constructed using the neighbor-joining analysis algorism. All positions containing gaps and missing data were eliminated. The Nearest-Neighbor-Interchange maximum likelihood heuristic search option was used for tree inference. Bootstrap analysis was performed using 1,000 replicates and evolutionary distances were calculated using the maximum composite likelihood method (Tamura et al., 2004)
Pathogenicity test.
The pathogenicity potential of the fungal isolates was tested using a 7-day culture in MEA. The mycelial plugs of each fungal species, cut to a size of 5.0 mm, were inoculated into five seedlings of the E. camaldulensis host plants through small wounds (2 to 3 cm) drilled into the stems with a scalper, at a depth sufficient to expose the xy-lem, approximately 30 cm above the collar root. Inoculation was done by placing the mycelial plugs upside down into the wound and covering it with masking tape. Sterile MEA discs free of fungal samples were used to inoculate the corresponding controls for comparison. The inoculated seedlings were carefully examined for symptoms of canker disease. After 4 months, canker symptoms were fully observed on the stems of the corresponding Eucalyptus seedlings. All resulting lesions were measured, sampled, cultured, re-isolated, and identified to confirm Koch's postulate (Pérez et al., 2009).
Statistical data analysis.
The data obtained were analyzed using SAS version 9.0 software procedures (SAS Institute Inc., 2011). Data on disease severity and incidence data were summarized, arranged, and expressed using descriptive statistics. The mean separation was determined by Duncan's multiple range test at P=0.01. Correlation and linear regression analysis were used to test the relationships between disease parameters and study sites and altitudes. Evolutionary history and phylogenetic analysis were performed using MEGA version 10 software (Kumar et al., 2018).
Results
Assessment of survey fields, disease symptoms, incidence, and severity.
The fields being studied were domi-nated by E. camaldulensis grown at their respective altitudes and often in non-agricultural rocky areas with infertile and compacted soils. Stem and branch canker diseases were observed, widely incident and severe in all plantation fields (Fig. 2). The E. camaldulensis plants were found to be relatively stressed in low-altitude areas. The minimum and maximum altitudes of the survey fields are 1,563 meters Abru (Abyotfire) and 2,254 meters Masken (Dirama) above sea level (Table 1). The highest, mean severity (90%) and incidence (100%) were recorded in the Amhara region, North Wollo, Abru district, and Abyot fire locality while the lowest average severity (45%) and incidence (66.2%) were recorded in the SNNP region, Gurage zone, Masken district, Dirama locality (Table 1). Stem and branch canker disease was recorded as more severe and incident at lower altitudes (Table 1).

Table 1.
Eucalyptus camaldulensis stem canker disease incidence and severity in 2019/2020 and 2020/2021
| Region | District | Locality | Host species | Altitude (m.a.s.l) | Mean±SD | |
|---|---|---|---|---|---|---|
| Incidence (%)a | Severity (%)a | |||||
| Oromia | Bedelle | Dabana | E. camaldulensis | 1,908 | 80±3.7 b | 55.8±0.8 c |
| Bedelle | Elike kerero | E. camaldulensis | 1,990 | 71±4.4 c | 55.6±2.5 c | |
| Jima | Dobi | E. camaldulensis | 1,676 | 82±0 b | 79.2±1.7 b | |
| SNNPR | Humbo | Gutoto larena | E. camaldulensis | 1,792 | 80.8±1.7 b | 55.4±2.6 c |
| Ana lamo | Laftolanka | E. camaldulensis | 2,100 | 65±0 c | 50±6.7 d | |
| Masken | Dirama | E. camaldulensis | 2,256 | 66.2±3.1 c | 45±4.2 e | |
| Amhara | Abru | Abyot fire | E. camaldulensis | 1,563 | 100±0 a | 90±3.5 a |
| Woreta | Woji | E. camaldulensis | 1,935 | 85.8±2.5 b | 54±0 c | |
| Mean | 78.8 | 60.6 | ||||
| P-value | <0.01 | <0.01 | ||||
| LSD | 6 | 2.5 | ||||
The occurrence and severity of stem and branch canker diseases were found to vary significantly (P<0.01) in the survey fields (Table 1). The results of the correlation test between the disease parameters (severity and incidence) of the canker diseases and the altitude of the study area show a significant negative relationship (r=‒0.82) at the 1% probability level. The result implies that the incidence and severity of stem canker diseases increase by 0.82 and 0.89 with a unit decrease in field altitude (Table 2).
Table 2.
Pearson's correlation (r), regression analysis using a linear model between the incidence and severity of stem canker disease with altitude and study locations
The result of the regression analysis using a linear model that relates the locations of the study areas as independent variables with the disease parameters as dependent variables show variables such as altitude in the surveyed areas influence (P<0.01) the incidence and severity of the disease with adjusted R2=0.87 and R2=0.99, respectively (Table 2). The result implies that environmental variables in the plantation fields have a strong impact on the significant variations in disease parameters in the study areas.
Morphological characterization.
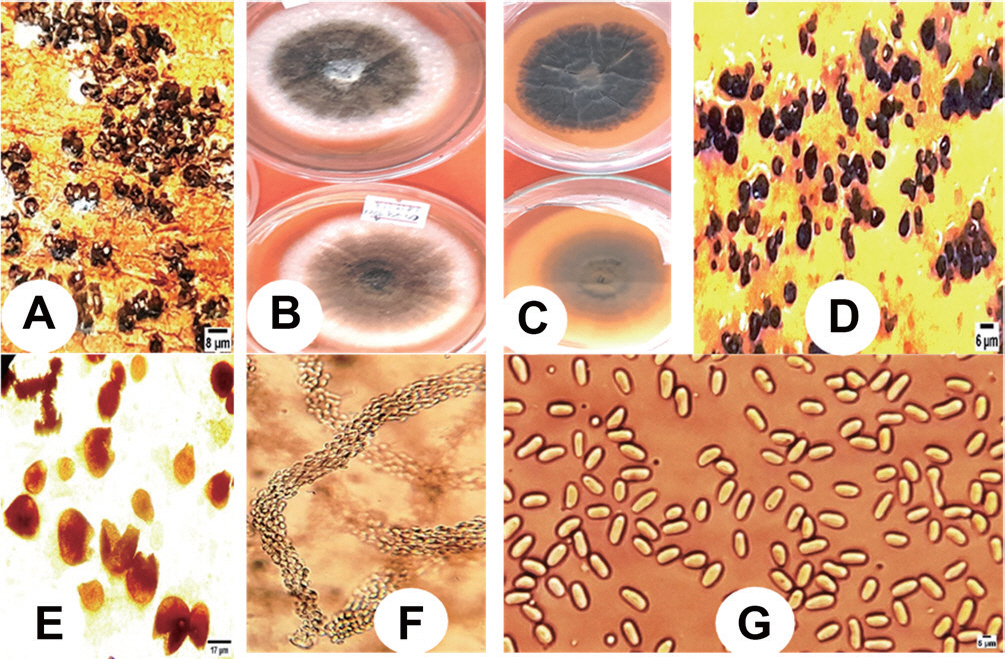
A total of 400 (320 branches and 80 stems) diseased samples of E. camaldulensis were collected from the survey fields. The fungal cultures were subcultured using the single-hyphal transfer method and several pure fungal isolates were obtained. A total of 120 fungal species were recovered from the collected samples. Isolation and morphological characterization of the fungal isolates were done using colony color, shape, zo-nation, structures of conidiomata, ascospores, and conidia morphs like shape and septation as indicated by Gezahgne et al. (2003). The bryoid and dendritic crystals formations associated with cultures were also used for morphological study as presented by Noordeloos et al. (1953). Most of the isolates (80%) were found Botryosphaeria, while the remaining 40% were Aplosporella and Didymella species described in this study. The Botryosphaeria and Aplosporella species studied in the research were submitted for publication. Research reports indicate that Didymella fungal species are currently the cause of emerging tree diseases in the world and that is why due consideration was given to the study (Chen et al., 2015). The fungal isolates in this study were cultured on MEA and the morphological characteristics were studied for three weeks as described by Noordeloos et al. (1953). The results of the fungal morphological character study revealed that the fungal growth was slow and the average radial diameter was recorded 75‒85 mm after 2 weeks. The colony was pale olivaceous gray to gray olivaceous, zonate, and/or with sectors; aerial mycelium was compact, white to gray, and flocculent on the front and reverse sides. The bryoid and dendritic crystals associated with the olivaceous black centres were formed after one week on the MEA (Fig. 3). Micromorphological observations were made using an Olympus BX53 digital fluorescence microscope. Pycnidia were abundant in concentric zones after crystals 3 weeks, but also scattered in the aerial mycelium and agar; black, globose or irregular; ostiolate solitary or mixed, sphere to pyriform, apical openings, superficial, brown to black with variable shape and size (Fig. 3). It was found that the conidia were light, ovate, aseptate, variable, smooth, and with slightly point-shaped formations at the tip (Fig. 3). The observed morphological characteristics were consistent with the genera of Didymella. The result agrees with the morphological features reported by Ahmadpour et al. (2017), Aveskamp et al. (2010), and Chen et al. (2015). The identification of the isolates to the key species level was performed by sequence analysis and phylogenetic study (Fig. 4).

DNA analysis and phylogenetic characterization.
The species, host, origins, and GenBank accession numbers, strains used for the phyllo-genetic study are displayed in Table 3. The phylogenetic tree was generated using the analysis of maximum likelihood (ML) algorithm (Fig. 4). The topology of the phylogenetic tree showed consistent relationships between the supported clades. Tree topology and phylogenetic analysis revealed that the fungal isolates in this study were associated with Didymella species (Fig. 4). The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1,000 replicates) are shown next to the branches. The tree is drawn to scale, with lengths next to the branches. The evolutionary distances were calculated using the maximum composite likelihood method (Tamura et al., 2004). Fungal species with close overlapping morphs and known outgroups were also included in the phylogeny for the reliability of the identification. This analysis was performed using 23 nucleotide sequences, and evolutionary analysis was performed in MEGA 10 (Kumar et al., 2018).
The best-fitting ML nucleotide substitution model for phylogenetic analysis of three-locus and four-locus com-bined datasets were selected Tamura-Nei (TN93+G+I) and General Time Reversible (GTR+G+I), respectively (Tamura, 1992). The phylogenetic tree of the dataset based on the ML analysis is shown in Fig. 4. The topology of the phylogenetic tree showed consistent relationships among the strongly supported clades. The tree topology of the phylogenetic analysis revealed that the isolates under study were associated with the species D. coffeae-arabicae and D. pinodella. In the tree, the isolates in our study (E864010FR002, 10B03-1; E864010FR003, 10B03-2; E864010FR004, 10B03-3) and (Q203241044, G11042; Q203241045, G11043) generated supportive monophyletic clades of 91 and 95% confidence levels with a reference strain from GenBank, Didymella coffeae-arabicae (MH863293, CBS 123380), and D. pinodella (MT274580:18-484, B36) (Fig. 4).
Table 3.
Fungal species used in the phylogenetic study
| GenBank accession no.a | Species | Isolate | Host | Origin |
|---|---|---|---|---|
| E864010FR 002 | D. coffeae-arabicae | 10 B03-1 | E.camaldulensis | Ethiopia |
| E864010FR 003 | D. coffeae-arabicae | 10 B03-2 | E.camaldulensis | Ethiopia |
| E864010FR 004 | D. coffeae-arabicae | 10 B03-3 | E.camaldulensis | Ethiopia |
| Q203241044 | D. pinodella | G11 042 | E.camaldulensis | Ethiopia |
| Q203241045 | D. pinodella | G11 043 | E.camaldulensis | Ethiopia |
| MK089255 | D. pinodella | T34 | Pisum sativum L. | Colombia |
| MH863293 | D. coffeae-arabicae | CBS 123380 | Coffea-arabica | Ethiopia |
| MK089252 | D. pinodella | T34 | Pisum sativum L. | Colombia |
| MG805350 | D. pinodes | WHDJ3 | Pisum sativum L. | China |
| MH855724 | D. pinodella | CBS 114.36 | n/a | Czechoslovakia |
| KY977440 | D. pinodella | SC08 | Sophora moorcroftiana | China |
| KX685357 | D. pinodes | NRCAb789 | Corchorus olitorius | Egypt |
| MW945409 | D. pinodella | ATCC 38814 | Pisum sativum | Canada |
| EU167565 | D. pinodella | CBS 110.32 | n/a | n/a |
| MN853862 | D. pinodella | UASWS2122 | Sequoiadendron giganteum | Netherlands |
| FJ427051 | D. pinodella | CBS 318.90 | Pisum sativum | Netherlands |
| FJ427052 | D. pinodella | CBS 531.66 | Trifolium pratense | USA |
| GU237829 | Leptosphaerulina australis | CBS 317.83 | Eugenia aromatica | Indonesia |
| MT274580:18-484 | D. pinodella | B36 | n/a | n/a |
| MN447424 | D. pinodella | C22 ITS1 | n/a | n/a |
| MT856268 | D. pinodella | BRR6 | n/a | China |
Based on the ITS sequence alignment in the BLAST search, the Didymella isolates in this study were 100% matched in sequence with D. coffeae-arabicae (GenBank no. MH863293) while the other fungal sequence in this study showed 99.79% identity with reference Didymella species in the NCBI GenBank (GenBank no. MT274580:18-484) (Fig. 4).
Pathogenicity test.
In this study, both D. pinodella and D. coffeae-arabicae isolates in the study caused lesions (Fig. 5) a month after inoculation of E. camaldulensis. There were no significant differences in the length of the lesion caused between the two species (P>0.01) (Table 4). Samples were collected, cultured in MEA, isolated, morphological characteristics were compared, and the results were consistent with the morphologies of isolation obtained from field samples. After the full onset of canker symptoms, small pieces of necrotic samples taken from the edges of the lesions were surface sterilized, cultured on the MEA, re-isolated and compared with morphological features of cultures obtained from diseased E. camaldulensis samples in the survey fields. The results were found consistent with the morphologies of the isolates obtained from the surveyed fields.
Fig. 5
Some of the lesions formed by Didymella pinodella (Dp) and D. coffeae-arabicae (Dc) (top) a month after inoculation of Eucalyptus camaldulensis seedlings with corresponding controls (bot-tom).

Table 4.
Mean lesion length data from Eucalyptus camaldulensis seedlings stem artificially inoculated for pathogenicity with the Didymella fungi
| Host | Fungal species | Isolate | Lesion size, mean±SD (mm) |
|---|---|---|---|
| E. camaldulensis | D. pinodella | G11 042 | 9.6±0.5a |
| G11 043 | 9.2±0.2a | ||
| D. coffeae-arabi- cae | 10 B03-1 | 9.7±0.6a | |
| 10 B03-2 | 9.9±0.4a | ||
| 10 B03-3 | 9.5±0.4a | ||
| P-value | >0.33 | ||
| LSD (1%) | 1.3 |
Discussion
In this study, E. camaldulensis plants were assessed for the disease symptoms, the level of damage was evaluated, samples were collected, isolated, morphologically and mo-lecularly characterized, tested for pathogenicity, and phylogenetic analysis was performed using reference species of Didymella in the GenBank.
E. camaldulensis plants in the survey fields were severely affected by the disease and showed typical symptoms of stem and branch canker diseases. Disease incidence and severity varied significantly (P<0.01) with the locations. The result of the regression analysis also confirmed that 87% to 99% of the variation in the incidence and severity of the canker diseases at the study sites was due to the environmental variables in the plantation fields. The results show that the occurrence and intensity of the disease depend on environmental variables such as altitude in plantation fields.
Pearson correlations (r) between disease incidence and severity with altitude in the survey field were negatively cor-related. Therefore, stem and branch canker diseases occur more incidentally and are more severe at lower elevations. Research reports by Golani et al. (2016), Maid and Ratnam (2014), Slippers and Wingfield (2007), and Wingfield et al. (2008) explain that environmental variables such as altitude influence the precipitation patterns of a particular area, resulting in seasonal humidity and temperature variations that create suitable conditions for the growth, reproduction, spread, and aggressiveness of fungal pathogens.
The results of the macroscopic and microscopic studies indicate that there are no significant morphological differences among the isolates of Didymella species in the study.
The ITS sequence study and the phylogenetic tree generated in this study show that the D. pinodella species (GenBank nos. E84010FR002, E864010FR003, E864010FR004) in this study have a strong monophyletic association with D. coffe-ae-arabicae, which was identified and reported from Ethiopia by Vu et al. (2019), while the rest of Didymella isolates were clustered with D. pinodella, designated by GenBank no. MT274580:180-484. Based on the sequence and phylogenetic study, the fungal species were identified as D. coffeae-arabicae and D. pinodella. According to Chen et al. (2017) and Keirnan et al. (2021), the genus Didymella consists of 35 known species that are widely distributed in both ornamen-tals and woody plants. Ahmadpour et al. (2017) reported that Didymella species such as D. microchlamydospora cause dieback and stem necrosis diseases in plants. Aveskamp et al. (2009) also coined the association of fungal species with Eucalyptus plant leaves. According to Barilli et al. (2016) and Chen et al. (2015), the genus Didymella is widely distributed in the fields and ornamental crops and can occur as a sap-robic, endophyte, or pathogen, causing diseases of blight, dieback, and necrosis diseases.
The results of the pathogenicity test with the identified fungal species confirm that the fungi are pathogenic and cause Eucalyptus canker diseases.
In this study, diverse fungal species were isolated, but according to their culture morphology and the importance of associated diseases, fungi belonging to the genera Phoma, Fusarium, Nothophom a, Cytospora, and Diaporthe were screened, sequenced, and identified. Furthermore, endophytic fungi such as Alternaria spp., Cladosporium spp., Epicoccum spp., Penicillium spp., and Nigrospora spp. were identified along with the Didymella species described in this study. The result implies the association of diverse fungal biota, that can cause diseases under favourable climatic conditions. This result is in agreement with the report of Sánchez Márquez et al. (2010). This finding affirms that the fungal species are globally distributed, have a broad host range, and can cause disease in economically important plant species. This is the first report of the fungal species associated with Eucalyptus plant disease in Ethiopia.
Despite limited access to the use of polymorphic regions in the molecular study, the ITS rDNA sequence and phylogenetic analyses enabled us to identify fungal isolates associated with E. camaldulensis stem canker diseases.
This research brings an additional new report that Didymellaceae is one of the fungal species causing stem and branch canker diseases of Eucalyptus plants worldwide. The findings of this study suggest that various fungal species associated with plants and causing diseases are still undiscovered. The study also indicates that the Eucalyptus canker disease in Ethiopia is due to synergies of several fungi triggered by stress due to local environmental factors and global climate changes at the moment. Appropriate agricultural practices that incorporate an integrated disease management system are the best approach to control plant diseases associated with adverse environmental factors. Further country-wise tree diseases survey, sample collection, and detailed phylogenetic studies are needed to obtain general information on fungal species and their host ranges that will help to develop effective disease management options at the country level.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






