



Introduction
Oat (Avena sativa, Poaceae) is one of the world's most important cereal crops, following maize, rice, wheat, barley, and sorghum (Hackett, 2018), and even though oat contains much higher unsaturated fatty acid, mineral vitamin, and phytochemical contents than other cereal species (Rasane et al., 2015), the evaluation of its quality has mainly focused on its value as a feed crop. Interestingly, recent studies that have confirmed the nutritional values and advantages of oat for human health have led to increases in oat consumption in Korea (Bana┼ø et al., 2007; Gorash et al., 2017), and oat can also grow well in poor environments (e.g., cold and humid climates) and exhibits a lower demand for chemical fertilizers than other grain crops (Lee et al., 2019).
Barley yellow dwarf virus (BYDV, Luteoviridae), a group of related single-stranded RNA viruses (Luteovirus, Polerovirus, or unassigned), are the most economically important and widespread viral pathogen of graminaceous crops, affecting over 150 species in the Poaceae (Bekele et al., 2001). A previous study separated BYDV into two subgroups that cor-responded to differences in the region coding for an RNA-dependent RNA polymerase (ORF2) (Malmstrom and Shu, 2004), with BYDV subgroup I including BYDV-PAV, BYDV-MAV, BYDV-SGV, and BYDV-PAS and subgroup II containing BYDV-RPV and BYDV-RMV. Of the various strains, BYDV-PAV is assumed to be the most widespread and is the most well studied at the molecular level (Jaro┼Īov├Ī et al., 2013). As with other luteoviruses, BYDV can be transmitted by various aphid species, and there is no evidence of mechanical or vertical transmission. The aphids Rhopalosiphum padi and Sitobian avanae are the primary vectors of BYDV (Thackray et al., 2009), and increases in aphid populations have been associated with both increases in BYDV incidence and sub-sequent reductions in crop production. Even though BYDV infections can be asymptomatic (McKirdy et al., 2002; Perry et al., 2000), BYDV-infected oat plants generally exhibit stunting, reduced root growth, and leaf discoloration (reddening), and BYDV has been reported to cause yield losses of up to 25% in barley, 46% in wheat, and 15% in oat, with a global mean of 30% (Adhikari et al., 2020; Edwards et al., 2001; Ordon et al., 2009). Most of the BYDV-PAV genome is Ōł╝5,660 bp in length, with six open reading frames and four untranslated regions (Liu et al., 2007). In particular, ORF3 encodes the main components of the coat protein (CP) required for aphid transmission and systemic infection (Filichkin et al., 1994; Gildow, 1993; Khine et al., 2020; Mohan et al., 1995).
Because oat is generally considered less important than wheat, rice, and barley, in terms of both quantity and profit-ability, the crop has attracted considerably less agricultural and disease research. The present study aimed to evaluate the incidence and distribution of BYDV in the oat-growing areas of Korea and to perform a phylogenetic analysis of BYDV isolates from Korea.
Materials and Methods
Sample collection.
Surveys were conducted during the oat-growing season (March to June) of 2020. The collected samples have symptoms, including dwarf, and reddening (Fig. 1A). Leaf samples were collected from eight different geographical regions of Korea (Yeoncheon, Pocheon, Suwon, Iksan, Gimje, Jeongeup, Gangjin, and Haenam) (Fig. 1B). Three hundred twenty-two samples were collected randomly from the commercial cultivation farm (Table 1). In particular, Jeongeup and Gangjin were the significant commercial production sites; thus, we collected many samples compared to other regions. All plant leaves were stored at -80┬░ C until the RNA extractions used for diagnostic tests.

RNA extraction.
Total RNA was extracted from homogenized leaf tissue using TRIzol (Ambion, Austin, TX, USA). Subsequently, both the concentration and quality of the resulting RNA samples were evaluated using a Nanodrop spectrophotometer (BioDrop, Cambridge, UK), and the RNA samples were stored at -20┬░C for further analysis.
Reverse transcription-polymerase chain reaction.
The incidence of BYDV infection in the collected oat leaf samples was assessed using reverse transcription-polymerase chain reaction (RT-PCR) and primers previously developed for a multiplex PCR protocol (Malmstrom and Shu, 2004). It is the most commonly used PCR primer based on the protocol for BYDV diagnostics (Table 2). The first step of the multiplex RT-PCR was to differentiate the BYDV isolates into two subgroups, with subgroup I including BYDV-PAV, BYDV-MAV, BYDV-SGV, and BYDV-PAS and subgroup II including BYDV-RPV and BYDV-RMV. Because of sequence differences between subgroups I and II, the forward primer (Shu-F) was used for subgroup identification. The second step of the multiplex PCR was designed to distinguish strains within subgroups I and II. Total RNA from the oat leaf samples screened for BYDV using the SuPrimeScript RT-PCR Premix (GeNet Bio, Daejeon, Korea). The 20-┬Ąl multiplex reactions included 0.4 ┬ĄM Shu-F, 0.3 ┬ĄM Yan-R, 0.07 ┬ĄM (total) of mixed S2a-F and S2b-F, 0.1 ┬ĄM PAV-F, 0.3 ┬ĄM (total) of mixed SGV1-R and SGV2-R, and 0.05 ┬ĄM (total) of mixed PASF and PASR, and 4 ┬Ąl total RNA. The RT-PCR conditions were as follows: initial reverse transcription (50┬░C, 30 min); polymerase activation (95┬░C, 5 min); 35 cycles of denaturation (95┬░C, 30 sec), annealing (55┬░C, 45 sec), and extension (72┬░C, 1 min); and a final extension (72┬░C, 7 min). All the RT-PCR products were analyzed using agarose gel (1.3% TBE) electrophoresis and UV visualization. Fragment sizes were determined by comparison to a 100-bp DNA marker.
Table┬Ā2.
Primers used to identify BYDV isolates from oat leaf samples from Korea
| Primer | Sequences 5ŌĆÖ-3ŌĆÖ | Specificity | Product size (bp) | Reference |
|---|---|---|---|---|
| Yan-R | TGTTGAGGAGTCTACCTATTTG | General | - | Malmstrom and Shu (2004) |
| Shu-F | TACGGTAAGTGCCCAACTCC | Subgroup I | With Yan-R: 830 | - |
| S2a-F | TCACCTTCGGGCCGTCTCTATCAG | Subgroup II (RPVs) | With Yan-R: 372 | - |
| S2b-F | TCACCTTCGGGGCGTCTCTTTCTG | Subgroup II (RMVs) | - | - |
| PAV-F | ACCTAGACGCGCAAATCAAA | PAVs | With Yan-R: 590 | - |
| SGV-R | ACATTTCTTCGTGTGTTGCG | SGVs | With Shu-F: 254 | - |
| ACATTTTTGCGTGCGTTGCG | - | - | - | |
| MAV2-F | AATAACCGCAGGAGAAATGG | MAVs | With Yan-R: 590 | - |
| PASF | GGAGACGACTGTGTCATCATCACTGAG | PASs | 448 | Laney et al. (2018) |
| PASR | TGTCGTTTGTGATAGGTGTCTCC | - | - | - |
PCR product sequencing and analysis.
Amplified DNA fragments were purified using a phenol/chloroform/ isoamyl alcohol solution. The purified DNA fragments were cloned into the T&A Easy T-vector (Yeastern Biotech, Taipei, Taiwan) and sequenced using M13F primers. Sequence identities were verified using BLAST to search the National Center for Biotechnology Information (NCBI) nucleotide database.
Sequence alignments and phylogenetic analy-ses.
The 23 BYDV CP sequences isolated from oat were compared to determine nucleotide identities and construct phylogenetic trees. The CP sequences of other BYDV isolates were downloaded from NCBI and included isolates from the USA (EF521840, EF521843), Australia (MK962883, X07653), Pakistan (KT252978, KT252976), Iran (KP771878), China (EU332333, AY855920), Estonia (MK012661), Germany (AJ810418), France (KC571999), and Japan (D85783). All 36 sequences were aligned and assembled using BioEdit version 7.0.5.3. Phylogenetic analysis of the BYDV-PAV CP sequence with other isolates was performed using the neighbor-joining method and Kimura 2-parameter method in MEGA 5.05 (Tamura et al., 2011), with 1,000 bootstrap replicates. Pairwise distances were calculated using SDT v1.2 (Muhire et al., 2014).
Results
BYDV incidence in oat in Korea.
Multiplex RT-PCR was performed to investigate the incidence of BYDV in oat in Korea. BYDV incidence was highest in Gimje (5/6, 83.3%), fol-lowed by Yeoncheon (9/12, 75%) and Suwon (11/16, 68.7%), and was lowest in Pocheon (0/4, 0%), Haenam (2/22, 9%). Intermediate levels of incidence were observed for samples from and Iksan (6/15), Jeongeup, and Gangjin. However, the incidence rates of samples from Jeongeup (n=128) and Gangjin (n=119), which were expected to be similar, were much higher in Jeongeup (48.4%) than Gangjin (24.3%). The overall incidence rate of BYDV infection was 38.7% (Table 3).
Table┬Ā3.
Distribution and incidence of BYDV subgroups in oat leaf samples collected from Korea in 2020
Identification of BYDV strains.
Multiplex RT-PCR was used to identify the specific BYDV stains present in the infected oat leaf samples. Among the infected samples (11/16) from Suwon, eight and three tested positive for BYDV-PAV and BYDV-RPV, respectively. The Jeongeup samples yielded the greatest number of BYDV strains, with BYDV-PAV detected in 51 samples, BYDV-RPV detected in eight samples, BYDV-SGV and BYDV-PAS each detected in one sample, and mixed infection (BYDV-PAV and BYDV-PAS) detected in one sample. Among the positive samples from Gangjin, 25 and four tested positive for BYDV-PAV and BYDV-SGV, respectively. Furthermore, 113 of the 125 BYDV-infected samples, including the sample with mixed infection, tested positive for Luteovirus, whereas 11 oat samples tested positive for Polerovirus. Furthermore, the most prevalent BYDV strains were BYDV-PAV, which was detected in 90% of the 113 Luteovirus-infected samples, and BYDV-PRV, which was detected in all 11 Polerovirus-infected samples. Interestingly, BYDV-MAV and BYDV-RMV were not detected in any region (Table 3).
Phylogenetic analysis.
CP sequences from BYDV-PAV isolates from eight regions (Yeoncheon, Pocheon, Suwon, Iksan, Gimje, Jeongeup, Haenam, and Gangjin) in Korea were deposited in GenBank (Table 4) and subject to phylogenetic analysis, along with 12 CP sequences from other regions of the world.
Table┬Ā4.
Accession numbers of CP nucleotide sequences of BYDV isolates from oat leaf samples collected from Korea in 2020
The phylogenetic analysis of BYDV CP sequences yielded two clades. Most (17/23) of the Korean CP sequences (LC567412, LC567413, LC550015, LC550016, LC567423, LC567424, LC567420, LC4567416, LC550012, LC550013, LC550011, LC567421, LC550014, LC567418, LC567426, LC567417, and LC567419) formed a group with one of the Chinese CP sequences (AY855920), and the other six Korean CP sequences (LC567425, LC530629, LC567415, LC567411, LC567422, and LC567414) formed a group with the remaining sequences from other counties. Phylogenetic analysis of complete CP sequences indicated that the 23 BYDV isolates from Korea were more closely related to each other (Fig. 2A).
Fig.┬Ā2.
Comparison of complete coat protein (CP) nucleotide sequences from Barley yellow dwarf virus isolates. (A) Phylogenetic tree. (B) Pairwise comparison. Information on the accession number and geographical location.
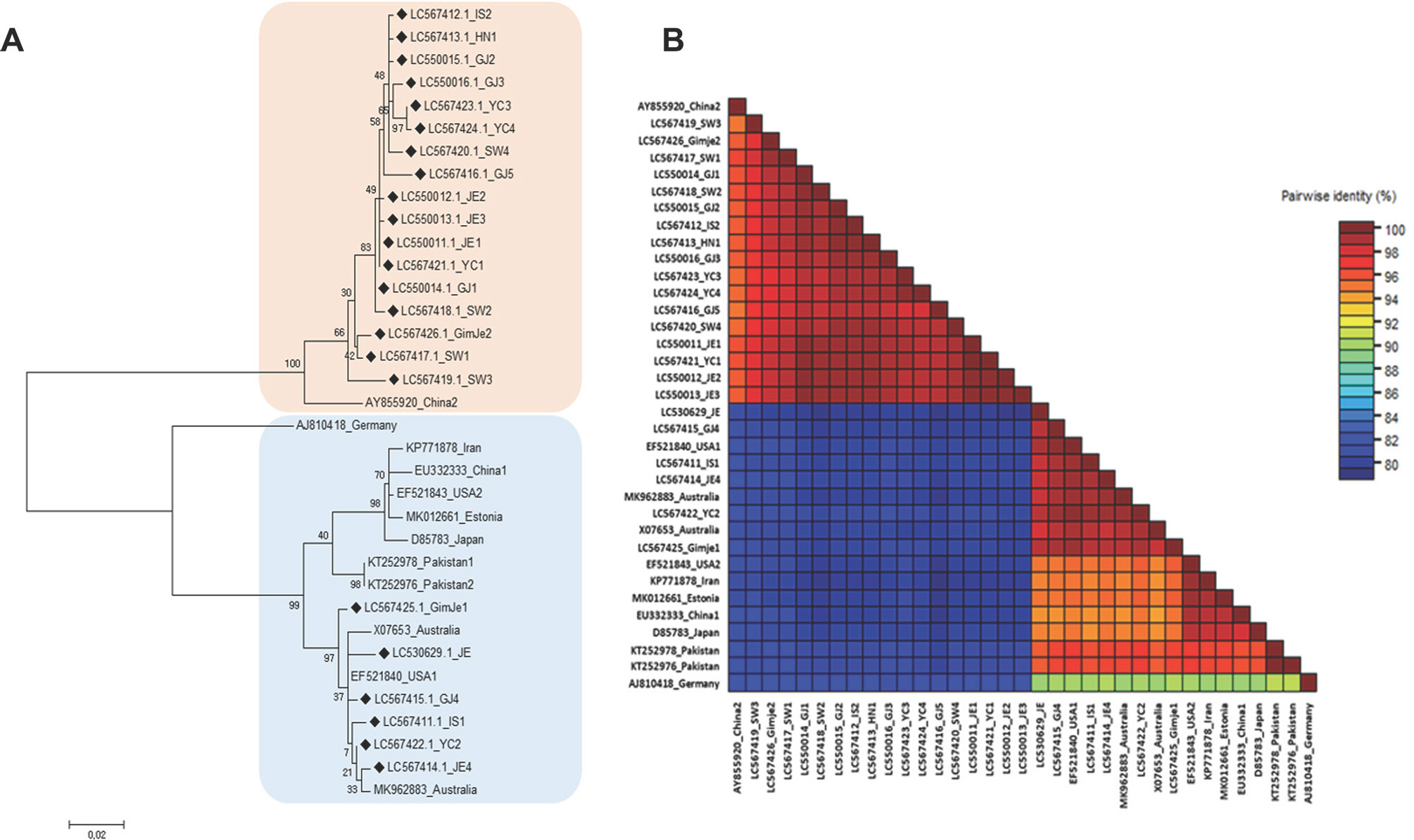
Pairwise comparison yielded similar results and allowed the separation of the sequences into two groups. The nucleotide sequence identity of the 17 Korean CP sequences in the first clade ranged from 97.0% to 99.7%, whereas those in the second clade ranged from 98.7% to 99.5%. The lowest sequence identity (79.5%) among the 23 Korean sequences was observed between JE1 (LC530629) and (LC567418) (Fig. 2B).
Discussion
Even though oat is one of the world's most widely cultivated cereal crops, its production has, until recently, focused more on forage than on food products (Capstaff and Miller, 2018). However, increase in public awareness of the crop's health benefits has resulted in rapid increases in oat consumption around the world (Rasane et al., 2015). In Korea, both the importation and domestic production of oats have increased. Oat is grown in several areas in Korea, including Gangjin (Jeollanam-do) and Jeongeup (Jeollabuk-do). BYDV represents a significant threat to cereal crop production, and although BYDV outbreaks in barley and wheat in Korea have been reported, current research regarding BYDV in oats is insufficient (Jo et al., 2018; Woo et al., 2001). Increasing global temperatures will likely promote increases in aphid populations (Ju et al., 2015), thereby increasing the occurrence of BYDV outbreaks and, thus, the resulting damage to affected crops. Therefore, investigation of the distribution of BYDV in Korea's domestic oat industry is critical.
In the present study, samples of symptomatic oat plants were collected from eight commercial oat fields across Korea, with most samples collected from Gangjin and Jeongeup, and multiplex RT-PCR was used to detect and identify BYDV strains in the samples. The results of the study suggest that sampling based on visual symptoms is ineffective. Indeed, symptoms that are similar to those of BYDV infection can also be caused by other diseases and physiological disorders, including nutrient stress, drought, and cold weather. In addition, the incidence of BYDV varied among the districts, from 0% (Pocheon) to 83% (Gimje), and BYDV-PAV was the most frequently detected strain, regardless of region, being detected in 33% of the 322 samples. Other BYDV strains (BYDV-SGV, BYDV-PAS, and BYDV-RPV) were also detected but were relatively rare, whereas others (BYDV-MAV and BYDV-RMV) were not detected in any region. Previous research has indicated that BYDV-PAV causes more severe symptoms and reductions in plant height and yield than other strains (Jaro┼Īov├Ī et al., 2013). Several strategies, including rapid detection, eradication, and breeding, have been developed for the control of viral crop pathogens. However, because BYDV incidence and damage will likely be promoted by climate change, due to the effects of increasing temperature on insect vectors, it will be increasingly important to continuously monitor BYDV and aphid outbreaks around the world.
In addition, BYDV-PAV isolates from different regions of Korea yielded divergent CP sequences. More specifically, the BYDV-PAV isolates were separated into two groups, which each contained closely related isolates. However, the overall sequence similarity of the 23 Korean BYDV CP sequences ranged from 79.5% to 99.7%. Interestingly, the Korean isolates were also more closely related to isolates from China and Australia than to those from other countries. This characterization and analysis provide insight into BYDV population dynamics and evolution.
The present study examined the status of BYDV in eight major major oat-producing areas in Korea and detected BYDV in 38.7% of collected samples using multiplex RT-PCR. However, the absence of BYDV in many symptomatic samples suggests that further research into other viral cereal crop pathogens is needed. The findings of the present study are expected to provide a basis for BYDV prevention, and continuous investigation is needed into BYDV transmission and interactions of the virus with vector insects.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






