



서론
인삼 (Panax ginseng Meyer, Araliaceae)은 한국, 일본, 중국을 비롯한 동아시아에서 수천 년간 재배되고 있는 중요한 의학용 작물이다(Park 등, 2012). 인삼은 뛰어난 약리 효과를 갖는 진세노사이드(ginsenosides)와 페놀성 화합물을 생성하며 약리 효과를 높이기 위해 4-6년의 생육기간이 필요하다(Chung 등, 2004). 다년의 생육기간 중, 인삼은 Botrytis cinerea, Cylindrocarpon destructans, Pythium spp., Rhizoctonia solani, Alternaria panax, Fusarium solani를 포함한 다양한 병원균의 위협에 노출되며 이러한 병원균은 인삼의 안정적 재배를 위협하는 주요 위험요인이다(Yu와 Ohh, 1993). 특히, C. destructans와 F. solani는 인삼뿌리썩음병을 일으켜 인삼연작을 방해하는 주요 원인이며, 토양과 이병식물조직에 후벽포자를 생성하여 불리한 환경 속에서도 10년 이상 생존할 수 있다(Kang 등, 2007).
연작장해를 경감하기 위해 다양한 경종적, 화학적, 생물적 방법이 도입되었다. 대표적인 경종적 방법은 4-5년 동안 벼를 재배한 곳에 인삼을 식재하는 것이다(Jo 등, 1996). 하지만 인삼을 이식 후 4-5년이 지나면 토양에 C. destructans의 밀도가 현저히 증가하고 인삼뿌리썩음병이 빈번히 발생하여 큰 피해를 양산한다(Park 등, 1997). 화학적 방제는 현재까지 가장 많이 사용되는 효과적인 방법이나, 살균제의 지속적 사용은 저항성 균주의 등장, 환경오염, 약해와 잔류농약 등의 문제를 야기한다(Handelsman과 Stabb, 1996; Hass와 Defago, 2005). 이러한 문제를 극복하기 위한 대안으로 Bacillus 속 세균을 활용한 생물적 방제 연구가 진행되고 있다(Kim 등 2012a; Kim 등, 2017; Song 등, 2014). Bacillus 속 세균은 항균 효과를 갖는 다양한 이차대사산물을 생성하는데, 대표적으로 cyclic lipopeptides (bacillomycin D, fengycin, iturin, surfactin), dipeptide (bacilysin), siderophore (bacillibactin), polyketides (bacillaene, difficidin, macrolactin)이 있다(Lee 등, 2012). 이와 같은 이차대사산물은 미생물 포자 발아와 생장을 억제하기 때문에 인삼을 비롯한 다양한 작물의 병원균에 대한 생물적 방제제로 활용되고 있다.
식물 내생균은 식물 생장 촉진, 생물·환경적 스트레스에 대한 내성, 이차대사산물 증진 등에 영향을 준다(Ryan 등, 2008). 인삼 내생세균이 인삼의 이차대사산물인 진세노사이드의 생성량을 증대시키고 인삼생장을 촉진시킬 수 있는 것으로 알려져 있다(Song 등, 2017). 또한 내생곰팡이 Trichoderma citrinoviride와 내생세균 Burkholderia stabilis는 인삼 병원균 생장을 억제하는 것으로 알려졌다(Kim 등, 2019; Park 등, 2019). 따라서 건강한 인삼으로부터 내생균을 분리하고 인삼뿌리썩음병원균에 대한 길항작용과 병발생에 미치는 영향을 분석하여 인삼뿌리썩음병을 억제하고 방제할 수 있는 미생물 후보군을 찾을 수 있을 것이다.
본 연구는 인삼뿌리썩음병의 원인균인 C. destructans와 F. solani의 생장을 억제하는 인삼 내생균을 찾아 이를 생물적 방제제로 활용하고자 수행되었다. 이를 위해 건강한 인삼에서 내생균을 분리하여 C. destructans와 F. solani 생장을 억제하는 세균을 선발하였다. 또한 선발된 세균 중에서 인삼뿌리썩음병원균이 생성하는 독소인 라디시콜(radicicol)에 대한 저항성을 갖는 내생균을 확인하였다. 선발 균주를 동정하고, in vitro 및 in vivo 조건에서 인삼뿌리썩음병원균에 대한 항균효과를 확인하는 실험을 수행했다. 또한, 항균활성 기작을 이해하기 위해 유전자 수준에서 항균물질 생합성을 조사했다. 본 연구 결과는 인삼내생균을 활용하여 인삼생육 전반에 인삼뿌리썩음병을 억제할 수 있는 미생물 제형 개발로 이어질 수 있을 것으로 기대한다.
재료 및 방법
인삼내생균 분리 및 인삼 병원성 진균 준비.
인삼 내생균을 분리하기 위해 인삼뿌리썩음병징을 보이지 않는 건강한 인삼을 두께 1 mm 절편으로 자른 후, 70% ethanol에서 1분, 5% sodium hypochlorite에서 3분간 표면살균 하였다. 표면살균한 인삼절편을 멸균수로 씻은 후, Luria-Bertani (LB) 고체배지에 치상하여, 25° C에서 2일간 배양하였다. 배양 중에 자라 나온 세균을 LB 고체배지로 옮겨 24시간 배양 후, 선상도말평판배양법(streak plate culture technique)으로 순수 균락을 확보하였다. 내생균의 장기 보관을 위해 40% glycerol 용액에 분리된 내생균을 넣은 후 -80° C에서 보관하였다. 인삼병원성 곰팡이 C. destructans 14seo 01-01 (CD-1), C. destructans KACC 41077 (CD-2), F. solani, Sclerotinia sclerotiorum KACC 40172, A. panax, B. cinerea를 감자한천배지(potato dextrose agar, PDA)에서 배양하였다(Kang 등, 2014).
대치배양.
분리된 내생균의 여러 가지 인삼병원균에 대한 균사생장 억제효과를 검정하기 위해 대치배양(dual culture) 실험을 수행하였다. 5가지 인삼병원균을 PDA 배지에 접종하고, CD-1, S. sclerotiorum은 20° C, F. solani, A. panax, B. cinerea는 25° C에서 5일간 배양하였다. 분리된 내생세균은 25° C, LB 고체배지에서 2일간 배양하였다. PDA에서 자란 곰팡이 균체를 지름 4 mm로 떼어내 새롭게 준비된 PDA 한편에 접종하고 4 cm 떨어진 맞은편에 내생세균을 접종하였다. CD-1, S. sclerotiorum은 20° C, F. solani, A. panax, B. cinerea는 25° C에서 6일간 배양한 후, 곰팡이 균사생장 억제 정도를 확인하였다. 대조군은 내생세균을 접종하지 않고 인삼병원균만 접종하였다.
포자발아 및 균사생장 억제효과 검정.
곰팡이포자 발아 억제 효과를 검정하기 위해 CD-1을 PDA에 접종하여 20oC, 암조건에서 7일간 배양했으며, F. solani는 carboxyl methyl cellulose에서 25° C, 200 rpm조건에서 4일간 배양하여 포자생성을 유도하였다. 내생세균은 brain heart infusion (BHI) 액체배지에서 28° C, 200 rpm 조건에서 7일간 배양 후, 13,000 rpm 10분간 원심분리 후 상등액을 0.2 μ m syringe filter (Corning Inc., Corning, NY, USA)에 통과시켜 무균의 배양여액을 확보하였다. 내생세균 배양여액이 곰팡이 포자 발아에 미치는 영향을 확인하기 위해 10 ml potato dextrose broth에 10 ml의 내생세균 배양여액을 처리하고 포자 농도가 1×106 conidia/ml이 되도록 접종하였다. 처리 후 CD-1은 20° C, F. solani는 25° C에서 200 rpm으로 배양하면서 4시간 간격으로 포자 발아율을 확인하였다. 내생세균 배양여액이 곰팡이 균사생장에 미치는 영향은 Kim 등(2017)이 제안했듯이 400 μ l의 배양여액을 PDA에 도말 후, 배지 가운데 CD-1와 F. solani 균체를 접종하고 20° C, 25° C에서 5일간 키운 뒤 처리군과 무처리군의 균총의 길이를 비교하였다.
Radicicol 저항성 검정.
Kang 등(2014)이 제안한 방법으로 분리된 인삼내생세균의 라디시콜 저항성을 확인하였다. 고압살균한 PDA를 50° C 정도로 식힌 후에 메탄올에 녹인 라디시콜을 50 mg/l 농도가 되도록 첨가하였다. 내생세균은 LB 고체배지에서 2일간 키운 후, 루프를 이용하여 단일 균총을 선상도말하였다. 배지를 25° C에서 배양하면서 라디시콜 처리군과 무처리군 간의 세균 생장을 관찰하였다.
분리균주 동정.
분리된 균주 동정을 위해 세균 분자계통 분류에 사용되는 16S rRNA 유전자 염기서열과 gyrA 유전자의 염기 서열을 해독하여 분류학적 위치를 분석하였다. 이를 위해 분리된 균주를 LB 배지에 24시간 키운 뒤 DokDo-Prep Genomic DNA kit (Elpis-Biotech Inc., Daejeon, Korea)를 이용하여 DNA 를 분리하였다. 16S rRNA 증폭을 위해 27F (5'-AGAGTTTGATC-MTGGCTCAG-3')와 1492R (5'-GGYTACCTTGTTACGACTT-3'), gyrA 증폭을 위해 p-gyrA-f (5'-CAGTCAGGAAATGCGTAC-GTCCTT-3')와 p-gyrA-r (5'-CAAGGTAATGCTCCAGGCATT-GCT-3') 프라이머(primer)를 이용하였다(Chun과 Bae, 2000). 증폭된 산물을 pGEMT-easy vector (Promega, Madison, MI, USA)에 cloning 후, Escherichia coli DH5α에 형질전환시켰다. 형질전환의 유무는 선택배지와 제한효소 Eco RI을 이용하여 확인한 후, plasmid를 분리하였다. 염기서열 분석은 T7 (5'-AATAC-GACTCACTATAG-3')과 SP6 (5'-ATTTAGGTGACACTATAG-3') 프라이머를 이용하여 Macrogen Inc. (Seoul, Korea)에서 수행되었다. 얻어진 염기서열을 SeqMan (DNASTAR, Madison, WI, USA)을 이용하여 오류를 검정하고 연결하였다. 16S rRNA 염기서열은 EzTaxon (http://www.ezbiocloud.net/eztaxon)에서 얻었으며(Kim 등, 2012b) gyrA 유전자의 염기서열은 GenBank (http://www.ncbi.nlm.nih/genbank)에서 얻었다. 계통도 작성을 위해 MEGA X 프로그램의 ClustalW (Thompson 등, 1994)를 이용하여 염기서열을 검정하고 염기서열 길이를 맞추었다. 계통도 작성은 maximum likehood 방법을 사용하였으며, 계통도의 정확성을 1,000회 반복 bootstrapping을 통해 확인하였다.
인삼뿌리썩음병 방제 효과 검정.
4년근 인삼을 70% ethanol에서 1분, 5% sodium hypochlorite에서 3분간 표면살균하였다. 드릴을 이용하여 한 뿌리에 총 8개의 구멍(지름 1 mm)을 뚫고, 10 μ l 포자현탁액(1×105 conidia/ml)을 구멍에 접종하였다. 내생세균은 단일 균총을 BHI 액체배지에 접종하고 28° C, 200 rpm에서 2일간 배양하였다. 배양액은 원심분리하여 세균을 모은 뒤 멸균수에 OD600=0.5 되게 현탁하고, 10 μ l를 구멍에 접종하였다. 처리군에는 병원균 접종 하루 전에 인삼내생균을 접종 후에 병원균을 접종하였다. 습도 유지를 위해 플라스틱 통에 젖은 휴지를 깔고 휴지와 접촉하지 않게 접종된 인삼을 위치시켰다. 플라스틱 뚜껑을 닫아 밀폐시킨 후, 20° C와 25° C, 암조건에서 14일간 병진전을 확인하였다.
항균물질 생합성 유전자 확인.
분리된 인삼내생균이 알려진 항균물질 생합성 유전자를 갖는지 여부를 중합효소연쇄반응(polymerase chain reaction, PCR)을 이용하여 확인하였다. PCR은 0.25 µl TaKaRa ExTaq (Takara Bio Inc., Kusatsu, Japan), 5 µl 10× ExTaq buffer, 4 µl dNTP mixture, 각 1 µl 프라이머 pair (10 µM) (Table 1), 1 µl DNA, 37.75 µl 증류수를 포함한 튜브에서 진행되었다. PCR 조건은 94° C로 3분 초기 변성후, 94° C 1분, 60° C 30초, 72° C 1분을 한 사이클(cycle)로 35회 반복하고, 72° C에서 최종반응이 수행되었다. PCR 산물은 0.005% ethid-ium bromide가 포함된 1.2% agarose gel에서 전기영동(125 V, 35분)을 진행시킨 후, UV illuminator를 통해 확인되었다.
Table 1.
Primers used in this s study
| Antifungal metabolite | Primer name | Sequence (5’→3’) | Reference |
|---|---|---|---|
| Zwittermicin A | ZWITF2 | TTGGGAGAATATACAGCTCT | Athukorala et al. (2009) |
| ZWITR1 | GACCTTTTGAAATGGGCGTA | ||
| Bacillomycin D | BACC1F | GAAGGACACGGCAGAGAGTC | Athukorala et al. (2009) |
| BACC1R | CGCTGATGACTGTTCATGCT | ||
| Iturin A | ITUD1F | GATGCGATCTCCTTGGATGT | Athukorala et al. (2009) |
| ITUD1R | ATCGTCATGTGCTGCTTGAG | ||
| Bacilysin | BACD-F1 | AAAAACAGTATTGGTYATCGCTGA | Chung et al. (2004) |
| BACD-R1 | CCATGATGCCTTCKATRCTGAT | ||
| Surfactin | SUR3F | ACAGTATGGAGGCATGGTC | Athukorala et al. (2009) |
| SUR3R | TTCCGCCACTTTTTCAGTTT | ||
| Fengycin | FEND1F | TTTGGCAGCAGGAGAAGTTT | Athukorala et al. (2009) |
| FEND1R | GCTGTCCGTTCTGCTTTTTC |
결과 및 고찰
건강한 인삼조직으로부터 총 17개의 내생세균을 분리하였다. 분리된 내생세균을 인삼뿌리썩음병의 원인균인 C. destructans와 F. solani에 대치배양한 결과, 3개의 균주(CH-9, CH-13, CH-15)가 두 병원균의 균사 생장을 효과적으로 억제하였다(Fig. 1A). 3개의 균주 중에서 CH-15가 다른 두 균주에 비해 C. destructans가 생성하는 독소인 라디시콜에 대해 큰 저항성을 보였다(Fig. 1B). 최종 선발된 CH-15 균주는 국립농업과학원 미생물은행(Korean Agriculture Culture Collection, KACC)에 기탁되었다(KACC No. 21772).
Fig. 1.
Antifungal activity and radicicol resistance of bacteria isolated from healthy ginseng. (A) Antifungal activity of the bacterial strains, CH-9, CH-13, and CH-15, against ginseng root rot pathogens. CD, Cylindrocarpon destructans 14seo 01-01; FS, Fusarium solani. (B) Radicicol resistance of C. destructans KACC41077 and the three bacterial strains.

인삼뿌리썩음병 방제를 위한 미생물을 스크리닝할 때 고려해야 할 중요한 요소가 바로 라디시콜에 대한 미생물의 저항성 여부이다. 라디시콜은 세포성장, 분화, 생존에 관련된 다양한 단백질들의 안정화 및 활성 조절에 중요한 역할을 하는 heat shock protein 90의 정상적 기능을 방해하여 미생물 생장에 영향을 미친다(Schulte 등, 1999). 또한 인삼뿌리썩음병원균을 제외한 다른 병원성 미생물과 토양 미생물의 생장을 저해하여 인삼뿌리썩음병원균이 토양에서 우점하고 병을 일으키는데 중요한 역할을 한다(Kang 등, 2014; Li 등, 2022). 따라서 인삼뿌리썩음병원균 방제를 위한 미생물 제제의 라디시콜에 대한 저항성은 인삼뿌리썩음병원균과 경쟁하는 데 중요한 역할을 할 것이며 효과적 미생물 제제를 선발하는 데 반드시 고려되어야 할 것이다.
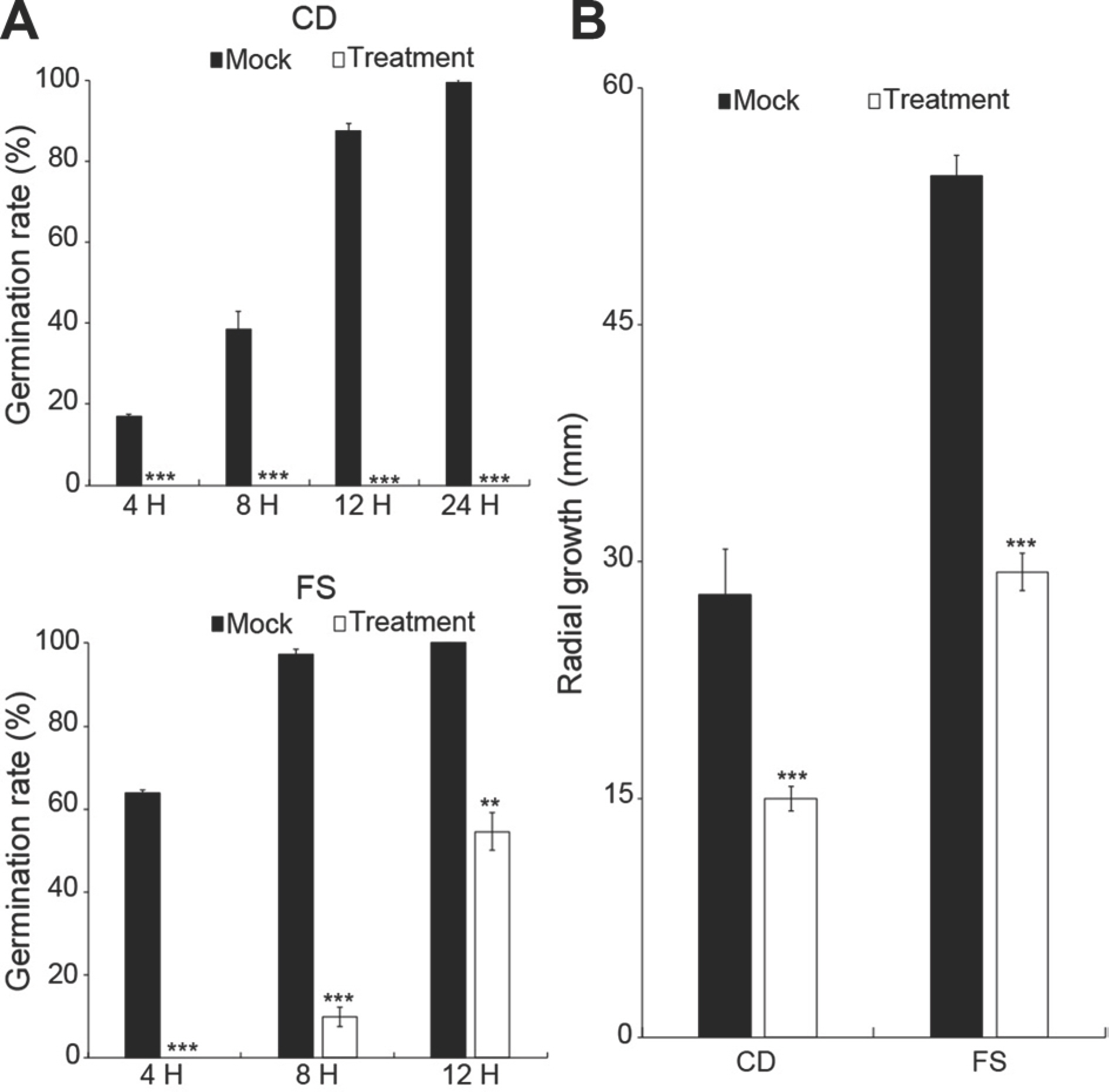
길항작용은 동일 환경에서 2종 이상의 미생물이 공존할 경 우 한 미생물이 다른 미생물의 생존과 증식을 위협하는 현상이며, 미생물이 생산하는 항생물질이 직접적인 원인인 경우가 많다(Kim 등, 2022). 이런 관점에서 길항세균 CH-15의 항균물질 분비여부를 확인하고자, 무균상태의 CH-15 배양여액의 병원균 생장과 포자 발아 억제 효능을 확인하였다. 그 결과, CH-15 배양여액은 C. destructans 포자 발아를 완전히 억제하였고, F. solani 포자 발아는 지연시켰다(Fig. 2A). 또한 CH-15 배양여액을 고체배지에 도말 후, 인삼뿌리썩음병원균을 접종했을 때, 무처리구에 비해 균사 생장이 현저하게 감소하였다(Fig. 2B).
Fig. 2.
Effect of CH-15 culture filtrate on spore germination (A) and mycelial growth (B) of ginseng root rot pathogens. Fungal spore germination rates of CD-1 was determined in 4, 8, 12, and 24 h after incubation at 20°C, and FS was determined in 4, 8, and 12 after incubation at 25°C. Mock did not contain culture filtrate. CD, Cylindrocarpon destructans 14seo 01-01; FS, Fusarium solani. Asterisks indicate P-value (∗∗ P<0.01,∗∗∗ P<0.001) after comparison with t-test, and data are presented as mean±standard deviation.
CH-15를 동정하기 위해 CH-15의 16S rRNA과 gyrA 유전자 염기서열을 분석한 결과, Bacillus velezensis B268의 16S rRNA 유전자 염기서열과 99.7% 일치하였고, Bacillus velezensis LA-BIM40의 gyrA 유전자 염기서열과 99.5% 일치하였다. 두 염기서열을 대상으로 선발 균주의 계통학적 분류 위치를 확인한 결과, CH-15는 B. velezensis와 같은 그룹에 위치하였다(Fig. 3).
Fig. 3.
Maximum likelihood phylogenetic tree based on partial 16S rRNA (A) and gyrA (B) gene sequences from Bacillus strains. Bootstrap frequencies are shown above the branches.

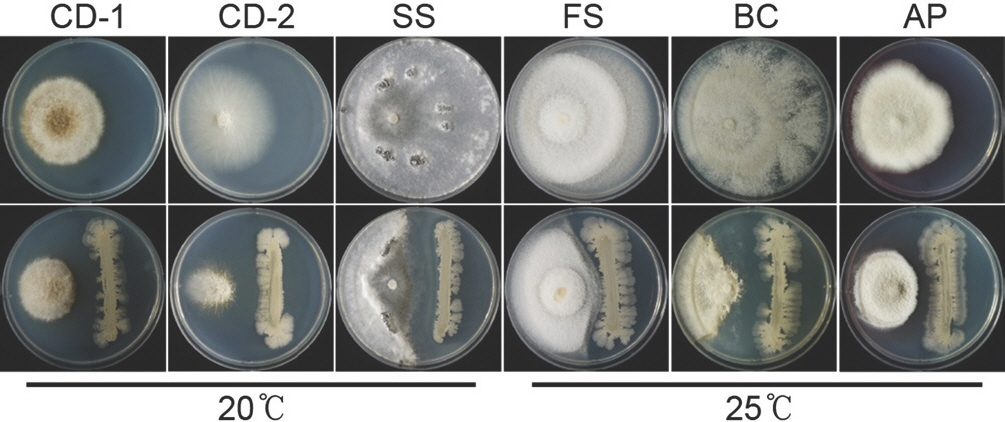
인삼뿌리에 구멍을 뚫고 C. destructans만을 접종했을 때 인삼 표면에 곰팡이 균사가 하얗게 자라나온 반면, C. destructans 를 접종하기 전에 CH-15를 처리하면 인삼표면에서 균사생장을 관찰할 수 없었다(Fig. 4A). 접종부위를 잘라 단면을 확인한 결과, C. destructans 단독 접종은 인삼 조직을 짙은 갈색으로 변화시킨 반면, C. destructans를 접종하기 전에 CH-15를 처리한 인삼 조직은 C. destructans를 접종하지 않은 무처리구와 비슷한 수준의 건강한 조직을 유지했다(Fig. 4A). Bacillus spp.는 항균효과를 갖는 다양한 물질을 생합성 하는 것으로 알려졌는데 그중 bacillomycin D, iturin A, bacilysin, surfactin은 Fusarium graminearum를 포함한 다양한 식물병원성 곰팡이에 항균 효과를 갖는 것으로 알려졌으며(Athukorala 등, 2009; Chung 등, 2008), PCR 결과는 CH-15가 이들을 생합성하는 유전자를 갖는다는 것을 보여준다(Fig. 4B). 비록 본 연구에서 항균활성물질을 규명하는 연구는 수행되지 않았으나 생합성 유전자가 존재하는 것으로 보아, CH-15가 보이는 항균 활성이 이러한 물질들에 의한 것일 가능성이 있다. 항균활성물질에 대한 정량분석과 정성분석을 통해 CH-15의 항균활성에 대한 기작을 밝히고 CH-15가 인삼내생균으로 존재할 때 식물체와 병원균에 미치는 영향을 이해한다면 인삼뿌리썩음병 방제를 위한 생물적 방제제로 개발 될 수 있을 것으로 기대한다. CH-15의 다양한 인삼병원균에 대한 항균활성을 확인한 결과, CH-15는 C. destructans 와 F. solani뿐만 아니라 S. sclerotiorum, B. cinerea, A. panax에도 항균활성을 나타냈다(Fig. 5). 이와 같은 결과는 인삼뿌리썩음병 뿐 아니라 다른 여러 가지 인삼병원균으로부터 인삼을 건강하게 재배하는데 있어서 CH-15의 활용 가능성을 보여준다.
Fig. 4.
Pathogenicity test of Panax ginseng and biosynthetic genes of antifungal metabolites. (A) Effect of CH-15 on ginseng root rot caused by Cylindrocarpon destructans 14seo 01-01. The ginseng 1-4 were inoculated with distilled water, CH-15, C. destructans, and both CH-15 and C. destructans 14seo 01-01, respectively. (B) The presence or absence of biosynthetic genes of antifungal metabo-lites in CH-15. L1, 100 bp DNA ladder; L2, zwiA (779 bp); L3, BmyA (875 bp); L4, ituA (647 bp); L5, bacD (749 bp); L6, srfA (441 bp); L7, fenD (964 bp); L8, 16S rRNA (1,511 bp); L9, λ DNA/ Hind III marker.
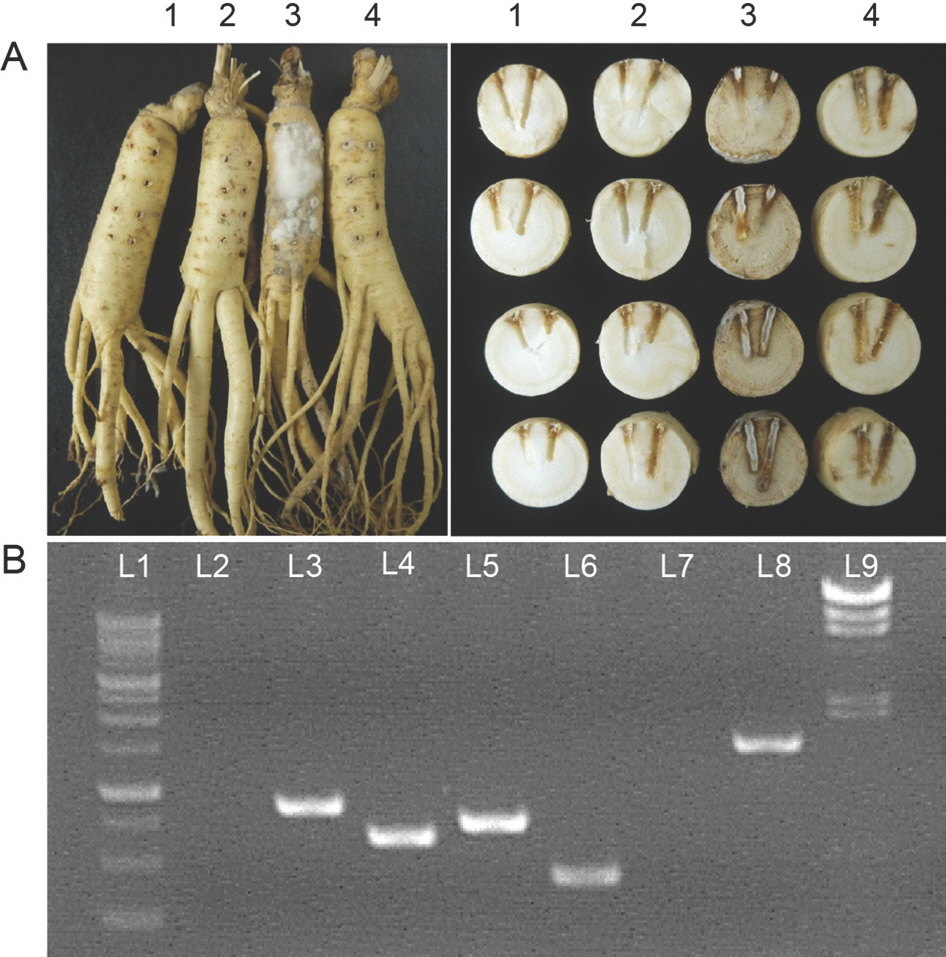
Fig. 5.
Antagonistic activity of CH-15 against ginseng pathogens. CD-1, Cylindrocarpon destructans 14seo 01-01; CD-2, C. destructans KACC 41077; SS, Sclerotinia sclerotiorum; FS, Fusarium solani; BC, Botrytis cinerea; AP, Alternaria panax.
본 연구는 건강한 인삼으로부터 분리한 미생물이 인삼뿌리썩음병원균과 다른 병원성 곰팡이에 대한 항균활성을 갖는다는 것을 보여준다. CH-15 균주의 사전 처리는 C. destructans의 균사생장을 억제할 뿐 아니라 병진전을 효과적으로 억제하였다. 또한 CH-15 균주는 C. destructans가 합성하는 독소인 라디시콜에 대해 저항성을 갖는다. 이는 CH-15 균주가 인삼뿌리썩음병원균과 같은 공간에 존재할 때 라디시콜에 영향 받지 않고 길항효과를 지속할 수 있을 것으로 기대한다. 특히, CH-15가 인삼내생균이기 때문에 인삼종자 발아 또는 유묘시에 처리하여 인삼에서의 내생을 유도하는 방법을 연구한다면, 인삼 생육 전주기에 걸쳐 발생하는 인삼뿌리썩음병을 억제하는 이상적인 생물적 방제 기술을 완성할 수 있다. 향후 연구에서 미생물 제형 개발, 인삼내생유도 및 기작, 내생균으로서의 인삼병원균 방제효과를 검정하는 연구를 거쳐 실효성을 확인한다면 인삼의 안정적 생산에 크게 기여할 수 있을 것으로 판단된다.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






