


ĒØ░Ļ░ĆļŻ©ļ│æ(Powdery mildew)ņØĆ ņśżņØ┤(Cucumis sativus)ļź╝ ĒżĒĢ©ĒĢśļŖö ļ¬©ļōĀ ļ░ĢĻ│╝(Cucurbitaceae) ņ×æļ¼╝ņŚÉņä£ Ļ░Ćņן ņŗ¼Ļ░üĒĢ£ Ēö╝ĒĢ┤ļź╝ ņØ╝ņ£╝ĒéżļŖö ļ│æ ņżæ ĒĢśļéśņØ┤ļŗż. ņśżņØ┤ ĒØ░Ļ░ĆļŻ©ļ│æņŚÉ Ļ░ÉņŚ╝ļÉśļ®┤ ņśżņØ┤ņØś ņ×ÄĻ│╝ ņżäĻĖ░ļź╝ ņżæņŗ¼ņ£╝ļĪ£ ĒØ░ņāē ļČäļ¦É ĒśĢĒā£ņØś ĻĘĀņé¼ņ▓┤ņÖĆ ļČäņāØĒżņ×ÉĻ░Ć ņāØņä▒ļÉśĻ│Ā, ņŗ¼ĒĢśĻ▓ī Ļ░ÉņŚ╝ļÉ£ ņ×ÄņØĆ Ļ┤æĒĢ®ņä▒ņØ┤ Ļ░ÉņåīĒĢśĻ│Ā ĻĮāĻ│╝ Ļ│╝ņŗżņØś ņāØņä▒ņŚÉļÅä ņśüĒ¢źņØä ņżĆļŗż(Rabelo ļō▒, 2017). Ļ░ÉņŚ╝ ņ┤łĻĖ░ ļŗ©Ļ│äņØś ĒØ░Ļ░ĆļŻ©ļ│æņØĆ ņŗØļ¼╝ ņ×Éņ▓┤ņŚÉ ņśüĒ¢źņØä ņŻ╝ņ¢┤ ņŗØļ¼╝ņØś Ļ┤┤ņé¼ļź╝ ņØ╝ņ£╝ĒéżĻ▒░ļéś ņłśĒÖĢļ¤ēņØä Ļ░Éņåīņŗ£ĒéżļŖö ļ░śļ®┤, Ļ░ÉņŚ╝ ĒøäĻĖ░ ļŗ©Ļ│äļŖö ņØ╝ļ░śņĀüņ£╝ļĪ£ Ļ│╝ņŗżņØś ĒÆłņ¦łņØä ņĀĆĒĢśņŗ£Ēé©ļŗż(Huang ļō▒, 2000). ņśżņØ┤ņŚÉ ĒØ░Ļ░ĆļŻ©ļ│æņØä ņØ╝ņ£╝ĒéżļŖö ļīĆĒæ£ņĀüņØĖ ļ│æņøÉĻĘĀņØĆ ļæÉ Ļ░Ćņ¦ĆņØś ņĀłļīĆĻĖ░ņāØĻĘĀ(obligate parasite) Golovinomyces cichoracearum (syn. Erysiphe cichoracearum)Ļ│╝ Podosphaera xanthii (syn. Sphaerotheca fuliginea)ņØ┤ļ®░(BertrandņÖĆ Pitrat, 1989; Hosoya ļō▒, 1999), Ēśäņ×¼ ĻĄŁļé┤ņŚÉņä£ ļŖö P. xanthiiĻ░Ć ņŻ╝ļĪ£ ĒØ░Ļ░ĆļŻ©ļ│æņØä ņØ╝ņ£╝ĒéżļŖö Ļ▓āņ£╝ļĪ£ ļ│┤Ļ│ĀļÉśņ¢┤ ņ׳ļŗż(Kim ļō▒, 2015). ņĄ£ĻĘ╝ ņŗ£ņäż ņ×¼ļ░░Ļ░Ć ņ”ØĻ░ĆĒĢśĻ│Ā ņś©ļé£ĒÖöĻ░Ć Ļ░ĆņåŹĒÖöļÉ©ņŚÉ ļö░ļØ╝ P. xanthiiņŚÉ ņØśĒĢ£ ļ│æ Ēö╝ĒĢ┤Ļ░Ć ņ”ØĻ░ĆĒĢśĻ│Ā ņ׳ļŖö ņČöņäĖņØ┤ļ®░ (Hong ļō▒, 2018; Tetteh ļō▒, 2013), ĒØ░Ļ░ĆļŻ©ļ│æ ņĀĆĒĢŁņä▒ ĒÆłņóģņØ┤ ņāłļĪŁĻ▓ī ņČ£ĒśäĒĢśļŖö raceņŚÉ ņØśĒĢ┤ ņØ┤ļ│æĒÖöļÉśļŖö ĒśäņāüņØ┤ ļ░śļ│ĄļÉśĻ│Ā ņ׳ļŗż. ļ│Ė ņŚ░ĻĄ¼ļŖö ĻĄŁļé┤ ņśżņØ┤ ĒØ░Ļ░ĆļŻ©ļ│æņØä ņØ╝ņ£╝ĒéżļŖö ņøÉņØĖĻĘĀņ£╝ļĪ£ P. xanthii isolatesņØä ļČäļ”¼ ļÅÖņĀĢĒĢśņśĆņ£╝ļ®░ ļ®£ļĪĀ ĒīÉļ│ä ņŗØļ¼╝ņØä ņØ┤ņÜ®ĒĢśņŚ¼ race 1ņÖĆ race 2ļĪ£ ĒīÉļ│äĒĢśņśĆļŗż. ļśÉĒĢ£, ĻĄŁļé┤ņŚÉņä£ ņāłļĪŁĻ▓ī ļ░£Ļ▓¼ļÉ£ race 2ņØś Ļ▓ĮņÜ░ ņČöĻ░ĆņĀüņØĖ ļ®£ļĪĀ ĒīÉļ│ä ĻĖ░ņŻ╝ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņĄ£ņóģņĀüņ£╝ļĪ£ race 2FļĪ£ Ļ▓░ņĀĢĒĢśņśĆļŗż.
ĻĄŁļé┤ ņśżņØ┤ ĒØ░Ļ░ĆļŻ©ļ│æ ņ×ÄņØś ņ▒äņ¦æ
ĻĄŁļé┤ņŚÉņä£ ļ░£ņāØĒĢśĻ│Ā ņ׳ļŖö ņśżņØ┤ ĒØ░Ļ░ĆļŻ©ļ│æĻĘĀ ņĪ░ņé¼ļź╝ ņ£äĒĢśņŚ¼ Ļ▓ĮĻĖ░ļÅä Ļ┤æņŻ╝(Gwangju), Ļ▓ĮĻĖ░ļÅä ņØ┤ņ▓£(Icheon), ņĀäļØ╝ļČüļÅä Ļ╣ĆņĀ£(Gimje), ņä£ņÜĖĒŖ╣ļ│äņŗ£(Seoul), Ļ▓Įņāüļé©ļÅä ļ░Ćņ¢æ(Miryang) ņ¦ĆņŚŁņØś ņ×¼ļ░░ļåŹĻ░ĆņŚÉņä£ ĒØ░Ļ░ĆļŻ©ļ│æņØ┤ ļ░£ņāØĒĢ£ ņśżņØ┤ ņ×ÄņØä ņ▒äņ¦æĒĢśņśĆļŗż(Fig. 1A, B). ņ▒äņ¦æĒĢ£ ņ×ÄņŚÉ ĒśĢņä▒ļÉ£ ĒØ░Ļ░ĆļŻ©ļ│æĻĘĀņØś ĻĘĀņŻ╝ļź╝ ļÅÖņĀĢĒĢśĻĖ░ ņ£äĒĢ┤ ņØ┤ļōżņØä ņØ┤ļ│æņä▒ņØĖ ņĪ░ņØĆļ░▒ļŗżļŗżĻĖ░ņśżņØ┤(FarmHannong, Seoul, Korea)ņØś ļ¢Īņ×Ä Ēś╣ņØĆ ļ│Ėņ×ÄņŚÉ ņ”ØņŗØĒĢśļŖö ļ░®ļ▓Ģ(Lee ļō▒, 2004)ņ£╝ļĪ£ ĒØ░Ļ░ĆļŻ©ļ│æĻĘĀņØä ņł£ņłś ļČäļ”¼ĒĢśņśĆļŗż.
ĒØ░Ļ░ĆļŻ©ļ│æĻĘĀņØś ņł£ņłś ļČäļ”¼ņÖĆ ņ”ØņŗØ
ņśżņØ┤ņÖĆ ļ®£ļĪĀņØĆ ņóģņ×É ņåīļÅģ Ēøä ļ®ĖĻĘĀļÉ£ ņ”ØļźśņłśņŚÉņä£ ļ░£ņĢäļź╝ ņ£ĀļÅäĒĢśņśĆĻ│Ā ļŗżĻ│Ą ĒöīļØ╝ņŖżĒŗ▒ ĒżĒŖĖņŚÉ ņøÉņśłņÜ®ņāüĒåĀ 2ĒśĖ(Seoul Bio, Gwangmyeong, Korea)ļź╝ ļäŻĻ│Ā ļ░£ņĢäļÉ£ ņóģņ×Éļź╝ ĒżĒŖĖļŗ╣ 1ļ”Įņö® ņŗ¼ņ¢┤ņŻ╝ņŚłļŗż. ļ¬©ļōĀ ņŗØļ¼╝ņØĆ 23┬░ 16ņŗ£Ļ░ä Ļ┤æņĪ░Ļ▒┤, 8ņŗ£Ļ░ä ņĢöņĪ░Ļ▒┤ņØ┤ ņ£Āņ¦ĆļÉśļŖö ņŗØļ¼╝ ņāØņןņāü Ēś╣ņØĆ ņäĖņóģļīĆĒĢÖĻĄÉ ņ║ĀĒŹ╝ņŖżņŚÉ ņ׳ļŖö ņ£Āļ”¼ņś©ņŗżņŚÉņä£ ņ×¼ļ░░ĒĢśņśĆļŗż. ņśżņØ┤ ĒØ░Ļ░ĆļŻ©ļ│æĻĘĀņØś ņł£ņłś ļČäļ”¼ņÖĆ ĒÜ©Ļ│╝ņĀüņØĖ ņ”ØņŗØņØä ņ£äĒĢśņŚ¼ ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö ņ×Ä ņĀłĒÄĖ(leaf disk)ņØä ņØ┤ņÜ®ĒĢ£ ļ░®ļ▓Ģ(Fanourakis, 1991; Lee ļō▒, 2004)ņØä ņØ╝ļČĆ ļ│ĆĒśĢĒĢśņŚ¼, Ļ░ÉņŚ╝ņ×Ä ņĀłĒÄĖņØä Ļ▒┤ņĀäņ×Ä Ēś╣ņØĆ Ļ▒┤ņĀäņ×Ä ņĀłĒÄĖĻ│╝ ņĀæņ┤ēņŗ£ņ╝£ ņĀæņóģņØä ņ£ĀļÅäĒĢśļŖö ŌĆśdisk-to-diskŌĆÖ ļ░®ļ▓ĢņØä ņé¼ņÜ®ĒĢśņśĆļŗż. Ļ▒┤ņĀäņ×Ä Ēś╣ņØĆ Ļ▒┤ņĀäņ×Ä ņĀłĒÄĖ(1.5├Ś1.5 cm)ņØä 70% ņŚÉĒāäņś¼ļĪ£ ņåīļÅģĒĢśĻ│Ā, ļ░░ņ¦Ć(25 mg/ml benzimidazol, 1% phytoagar) (Gafni ļō▒, 2015)Ļ░Ć ļČäņŻ╝ļÉ£ ĒÄśĒŖĖļ”¼ļööņē¼ņŚÉ ņ╣śņāüĒĢ£ Ēøä ņ×¼ļ░░ ļåŹĻ░ĆņŚÉņä£ ņ▒äņ¦æĒĢ£ ĒØ░Ļ░ĆļŻ©ļ│æ Ļ░ÉņŚ╝ņ×ÄņØś ņĀłĒÄĖ(0.8├Ś0.8 cm)ņØä Ļ▒┤ņĀäņ×Ä Ēś╣ņØĆ Ļ▒┤ņĀäņ×Ä ņĀłĒÄĖ ņ£äņŚÉ ņś¼ļĀżņŻ╝Ļ│Ā ļ░Ćļ┤ēĒĢśņśĆļŗż. ĒØ░Ļ░ĆļŻ©ļ│æĻĘĀņØĆ ļ░Ćļ┤ēļÉ£ ĒÄśĒŖĖļ”¼ļööņē¼ņŚÉņä£ 23┬░, 24ņŗ£Ļ░ä Ļ┤æņĪ░Ļ▒┤ņ£╝ļĪ£ ņĢĮ 10ņØ╝Ļ░ä ņ”ØņŗØĒĢśĻ│Ā, ņØ┤ņāüņØś ņ”ØņŗØņØä ņĄ£ņåī 3ĒÜī ņØ┤ņāü ļ░śļ│ĄĒĢśņŚ¼ ņł£ņłś ļČäļ”¼ĒĢśņśĆļŗż. Ļ▓ĮĻĖ░ļÅä Ļ┤æņŻ╝, Ļ▓ĮĻĖ░ļÅä ņØ┤ņ▓£, ņĀäļØ╝ļČüļÅä Ļ╣ĆņĀ£,ņä£ņÜĖĒŖ╣ļ│äņŗ£, Ļ▓Įņāüļé©ļÅä ļ░Ćņ¢æņŚÉņä£ ņłśņ¦æļÉ£ ĒØ░Ļ░ĆļŻ©ļ│æĻĘĀņØś ļČäļ”¼ ĻĘĀņŻ╝ļź╝ Ļ░üĻ░ü GW170710, IC190611, KI170723, SE180328, MI180427ļØ╝Ļ│Ā ļ¬ģļ¬ģĒĢśņśĆļŗż.
ĻĄŁļé┤ ĒØ░Ļ░ĆļŻ©ļ│æĻĘĀņØś ĒśĢĒā£ĒĢÖņĀü Ļ┤Ćņ░░Ļ│╝ ņŚ╝ĻĖ░ņä£ņŚ┤ ļČäņäØ
ļČäļ”¼ ĻĘĀņŻ╝ļōżņØś ĒśĢĒā£ņĀü ļČäņäØņØä ņ£äĒĢśņŚ¼ ņ”ØņŗØļÉ£ ĒØ░Ļ░ĆļŻ©ļ│æĻĘĀņØś ĻĘĀņé¼ņ▓┤ļź╝ ļ®ĖĻĘĀļÉ£ Ēīüņ£╝ļĪ£ ĻĖüņ¢┤ļé┤ņ¢┤ ļ®ĖĻĘĀ ņ”Øļźśņłś 200 ╬╝lļĪ£ ĒؼņäØĒĢśĻ│Ā Ļ┤æĒĢÖĒśäļ»ĖĻ▓Į(Eclipse Ti, Nikon, Tokyo, Japan)ņŚÉņä£ Ļ┤Ćņ░░ĒĢśņśĆļŗż. ĻĄŁļé┤ņŚÉņä£ ņ▒äņ¦æĒĢ£ ņśżņØ┤ ĒØ░Ļ░ĆļŻ©ļ│æņØś 5Ļ░Ćņ¦Ć ļČäļ”¼ ĻĘĀņŻ╝ļōżņØś(KI170723, GW170710, SE180328, MI180427 ļ░Å IC190611) Ēżņ×ÉļōżņØĆ ļ¼┤ņāē ĒāĆņøÉĒśĢņØś ĒśĢĒā£ĒĢÖņĀü ĒŖ╣ņ¦ĢņØ┤ P. xanthiiņÖĆ ļÅÖņØ╝ĒĢśņśĆļŗż(Fig. 1C). Ļ░ü ņ¦ĆņŚŁļ│ä ĒØ░Ļ░ĆļŻ©ļ│æĻĘĀņØś ļČäņ×ÉņāØļ¼╝ĒĢÖņĀü ĒŖ╣ņ¦ĢņØä ļ╣äĻĄÉĒĢśĻĖ░ ņ£äĒĢśņŚ¼ KI170723, GW170710, SE180328, MI180427 ļ░Å IC190611ņØś genomic DNAļź╝ cetyl trimethyl ammonium bromide (CTAB)ļ▓Ģ(Porebski ļō▒, 1997)ņØä ņØ┤ņÜ®ĒĢśņŚ¼ ņČöņČ£ĒĢśņśĆļŗż. ņäĀĒ¢ē ņŚ░ĻĄ¼(Pirondi ļō▒, 2015)ļź╝ ļö░ļØ╝ņä£ internal transcribed spacer (ITS) ņśüņŚŁņØä ITS1 (5ŌĆ▓-TCCGTAGGTGAACCTGCGG-3ŌĆ▓)Ļ│╝ ITS4 (5ŌĆ▓-TCCTCCGCTTATTGATATGC-3ŌĆ▓)ļź╝ ņé¼ņÜ®ĒĢśņŚ¼ ņżæĒĢ®ĒÜ©ņåīņŚ░ņćäļ░śņØæ(polymerase chain reaction)ņ£╝ļĪ£ ņ”ØĒÅŁņŗ£ņ╝░ļŗż. ņ”ØĒÅŁļÉ£ ņé░ļ¼╝ļōżņØś ņŚ╝ĻĖ░ņä£ņŚ┤ņØĆ BLAST search (https://blast.ncbi.nlm.nih.gov/Blast.cgi)ļź╝ ņØ┤ņÜ®ĒĢ£ ļ╣äĻĄÉ ļČäņäØ Ļ▓░Ļ│╝ GenBanKņŚÉ ļō▒ļĪØļÉ£ P. xanthiiņØś ITS (accession no. KJ698669.1)ņØś ņŚ╝ĻĖ░ņä£ņŚ┤Ļ│╝ 100% ņØ╝ņ╣śĒĢśņśĆļŗż(data not shown). ļö░ļØ╝ņä£, ņ▒äņ¦æļÉ£ ĒØ░Ļ░ĆļŻ©ļ│æĻĘĀļōżņØś ļČäļ”¼ĻĘĀņŻ╝ļōżņØĆ P. xanthiiņ×äņØä ņĢī ņłś ņ׳ņŚłļŗż.
ĒØ░Ļ░ĆļŻ©ļ│æĻĘĀ ļČäļ”¼ ĻĘĀņŻ╝ļōżņØś race ĒīÉļ│ä
ļ│æ ņĀĆĒĢŁņä▒ ĒÆłņóģ ņ£Īņä▒ņØä ņ£äĒĢ┤ņä£ļŖö ņĀĆĒĢŁņä▒ Ļ▓ĆņĀĢņŚÉ ņé¼ņÜ®ĒĢĀ ļ│æņøÉĻĘĀņØś race ĻĘ£ļ¬ģņØ┤ ņäĀĒ¢ēļÉśņ¢┤ņĢ╝ ĒĢśļŖöļŹ░, ņśżņØ┤ņŚÉņä£ ļ░£ļ│æĒĢśļŖö P. xanthii Ļ▓ĮņÜ░ ļŗżņ¢æĒĢ£ ņāØļ”¼ĒśĢ(physiological race)ļōżņØ┤ ļ│┤Ļ│ĀļÉśņ¢┤ ņ׳ņ£╝ļéś ņĀĢļ”ĮļÉśņ¢┤ ņ׳ņ¦Ć ņĢŖņĢä ņ£ĪņóģņŚÉ ņØ┤ņÜ®ĒĢśĻĖ░ ņ¢┤ļĀĄļŗż(Lee ļō▒, 2014). ļīĆņŗĀņŚÉ P. xanthii ļŖö ļ░ĢĻ│╝ ņ×æļ¼╝ņØĖ ļ®£ļĪĀ, ņłśļ░Ģ, ņśżņØ┤ļź╝ ļ¬©ļæÉ ĻĄÉņ░©Ļ░ÉņŚ╝ĒĢśĻĖ░ ļĢīļ¼ĖņŚÉ (Cohen ļō▒, 2000) ņŻ╝ļĪ£ ļ®£ļĪĀ ĒīÉļ│ä ĒÆłņóģņØä ņØ┤ņÜ®ĒĢśņŚ¼ raceļź╝ Ļ▓░ņĀĢĒĢśĻ│Ā ņ׳ļŗż(Kuzuya ļō▒, 2006). 1935ļģä ļ»ĖĻĄŁņŚÉņä£ ņ£Īņä▒ļÉ£ ļ®£ļĪĀ ĒØ░Ļ░ĆļŻ©ļ│æ ņĀĆĒĢŁņä▒ Ļ│äĒåĄņØĖ ŌĆśPMR 45ŌĆÖņŚÉ ļ░£ļ│æĒĢśļŖö ĻĘĀņØä race 1, ŌĆśPMR 45ŌĆÖņŚÉ ļ░£ļ│æĒĢśņ¦Ć ņĢŖļŖö ĻĘĀņØä race 2ļĪ£ ļČäļźśĒĢśņśĆļŗż(Jagger ļō▒, 1938). 1946ļģä ļ»ĖĻĄŁņŚÉņä£ ņ£Īņä▒ļÉ£ race 1Ļ│╝ race 2ņŚÉ ņĀĆĒĢŁņä▒ņØä Ļ░¢ļŖö ŌĆśPMR 5ŌĆÖļź╝ ņØ┤ļ│æĒÖö ņŗ£ĒéżļŖö ĻĘĀņØä race 3ņ£╝ļĪ£ ļČäļźśĒĢśņśĆļŗż(Pryor ļō▒, 1946). ņØ┤ļź╝ ĻĖ░ļ│Ėņ£╝ļĪ£ ĒĢśņŚ¼ P. xanthiiņØś race ĒīÉļ│äņØĆ ņØ╝ļ░śņĀüņ£╝ļĪ£ 4Ļ░Ćņ¦Ć ļ®£ļĪĀ ĒÆłņóģ(ŌĆśTopmarkŌĆÖ, ŌĆśPMR 45ŌĆÖ, ŌĆśPMR 6ŌĆÖ ļ░Å ŌĆśMR-1ŌĆÖ)ņØä ņØ┤ņÜ®ĒĢśņŚ¼ race 1, 2, 3ņ£╝ļĪ£ ĒīÉļ│äĒĢśĻ│Ā ņ׳ļŗż(Zitter et al., 1996) (Table 1). ļ│Ė ņŚ░ĻĄ¼ņØś race ĒīÉļ│äņØä ņ£äĒĢ┤ņä£ļŖö ņ£äņŚÉņä£ ņäżļ¬ģĒĢ£ ļ░öņÖĆ Ļ░ÖņØ┤ ņ×Ä ņĀłĒÄĖņØä ņØ┤ņÜ®ĒĢ£ ļ░®ļ▓Ģ(Fanourakis, 1991; Lee ļō▒, 2004)ņØä ņØ╝ļČĆ ļ│ĆĒśĢĒĢ£ ŌĆśdisk-to-diskŌĆÖļ▓ĢņØä ņØ┤ņÜ®ĒĢśņśĆļŗż. ņĀæņóģ Ēøä ņĢĮ 14ņØ╝ņŚÉ ņé¼ņ¦ä ņ┤¼ņśü Ēøä ImageJ ĒöäļĪ£ĻĘĖļש(ver. 1.53c, National Institutes of Health, Bethesda, MD, USA)ņØś ŌĆ£Color thresholdŌĆØ (https://imagej.nih.gov/ij/)ņØä ņØ┤ņÜ®ĒĢśņŚ¼ ņĀĢļ¤ēĒÖöĒĢśņśĆņ£╝ļ®░ Ļ░ÉņŚ╝ņ×Ä ņĀłĒÄĖņØś Ēü¼ĻĖ░ļź╝ ĻĖ░ņżĆņ£╝ļĪ£ Ļ▒┤ņĀäņ×ÄņØś ĒØ░Ļ░ĆļŻ©ļ│æ ļ│æļ░śļ®┤ņĀüļźĀ(%)ņØä Ļ│äņé░ĒĢśņśĆļŗż. ĒÅēĻĘĀ ļ░£ļ│æ ņĀĢļÅäĻ░Ć 10% ņØ┤ĒĢśņØś Ļ▓ĮņÜ░ņŚÉļŖö ņĀĆĒĢŁņä▒(resistant, R) 11%-30%ļŖö ņżæļÅäņĀĆĒĢŁņä▒(intermediate resistant, IR), 31% ņØ┤ņāüņØĆ Ļ░Éņłśņä▒(susceptible, S)ņ£╝ļĪ£ ĒīÉņĀĢĒĢśņśĆļŗż.
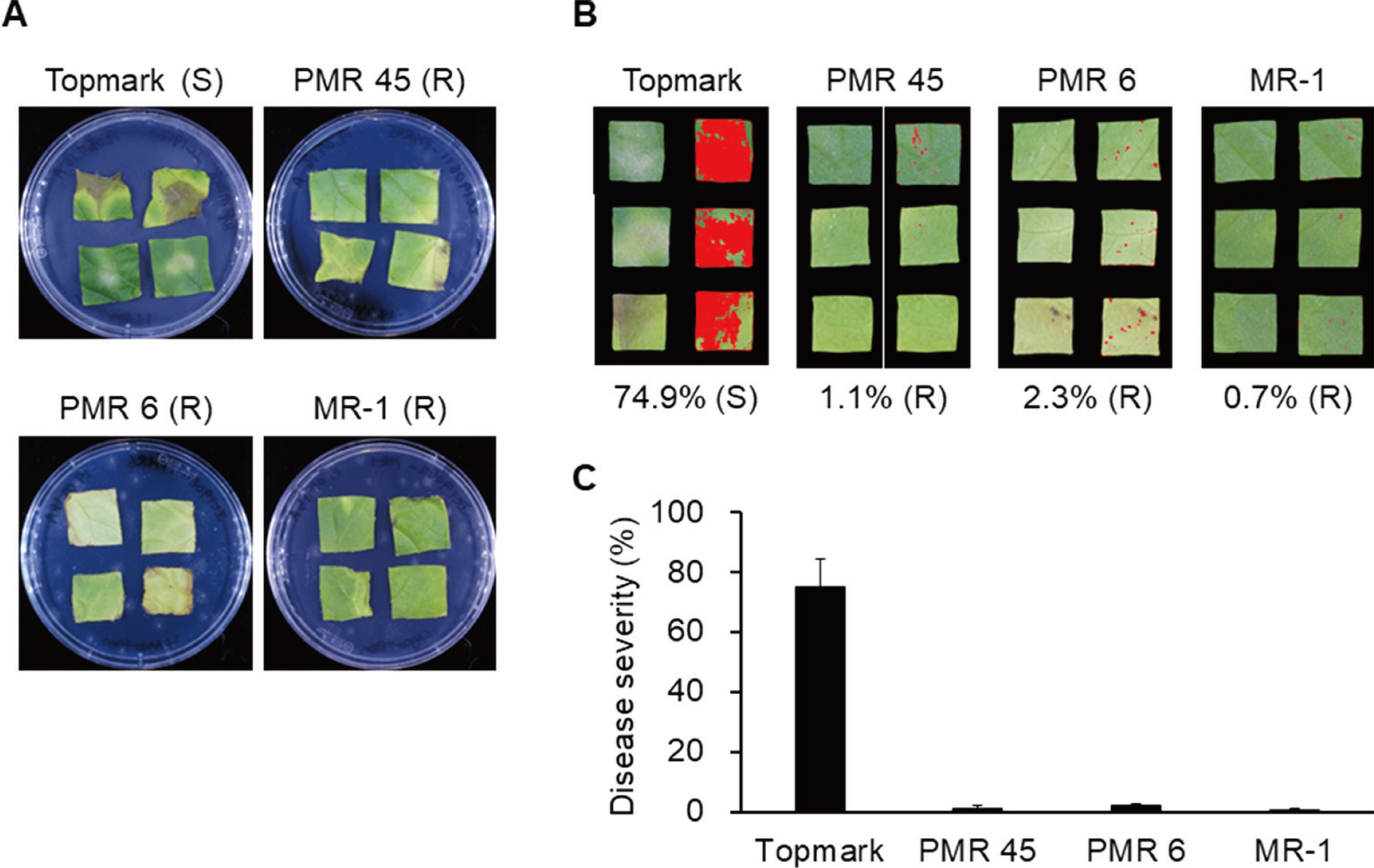
ļČäļ”¼ ĻĘĀņŻ╝ MI180427ņÖĆ IC190611ņØś raceļź╝ ĻĘ£ļ¬ģĒĢśĻĖ░ ņ£äĒĢ£ ņĀæņóģ Ļ▓░Ļ│╝, MI180427ļŖö ŌĆśTopmarkŌĆÖņŚÉņä£ļŖö 51.1%ļĪ£ Ļ░Éņłśņä▒(S)ņØä ļ│┤ņśĆņ£╝ļ®░, ŌĆśPMR 45ŌĆÖņŚÉņä£ 1.6%, ŌĆśPMR 6ŌĆÖņŚÉņä£ 0.6% ĻĘĖļ”¼Ļ│Ā ŌĆśMR-1ŌĆÖņŚÉņä£ 0.3%ņØś ļ░£ļ│æ ņĀĢļÅäļĪ£ ņĀĆĒĢŁņä▒(resistance, R)ņØä ļéśĒāĆļé┤ņŚłļŗż(Fig. 2). IC190611ļŖö ŌĆśTopmarkŌĆÖņŚÉņä£ 74.9% (S), ŌĆśPMR 45ŌĆÖņŚÉņä£ 1.1% (R), ŌĆśPMR 6ŌĆÖņŚÉņä£ 2.3% (R) ĻĘĖļ”¼Ļ│Ā ŌĆśMR-1ŌĆÖņŚÉņä£ 0.7% (R)ņØś ļ░£ļ│æ ņĀĢļÅäļź╝ ļéśĒāĆļé┤ņŚłļŗż(Fig. 3). ļö░ļØ╝ņä£, ļ░Ćņ¢æĻ│╝ ņØ┤ņ▓£ņŚÉņä£ ļČäļ”¼ĒĢ£ ĒØ░Ļ░ĆļŻ©ļ│æĻĘĀņØĆ Zitter ļō▒(1996)ņØś race ĒīÉļ│ä ĻĖ░ņżĆņŚÉ ļö░ļØ╝ race 1ņØ┤ļØ╝ļŖö Ļ▓āņØä ņĢī ņłś ņ׳ņŚłļŗż (Table 1). P. xanthiiņØś ņŚ¼ļ¤¼ race ņżæ race 1ņØĆ ļ»ĖĻĄŁ ļÅÖļČĆņ¦ĆņŚŁņŚÉņä£ Ļ░Ćņן ĒØöĒĢśĻ▓ī ņĪ┤ņ×¼ĒĢśļŖö ņŻ╝ņÜöĒĢ£ ĒØ░Ļ░ĆļŻ©ļ│æĻĘĀņØ┤ļ®░(Zitter et al., 1996), ĻĄŁļé┤ņŚÉņä£ļÅä ņÜ░ņĀÉĒĢśļŖö Ļ▓āņ£╝ļĪ£ ļ│┤Ļ│ĀļÉ£ raceņØ┤ļŗż(Kim ļō▒, 2016; Lee ļō▒, 2014).
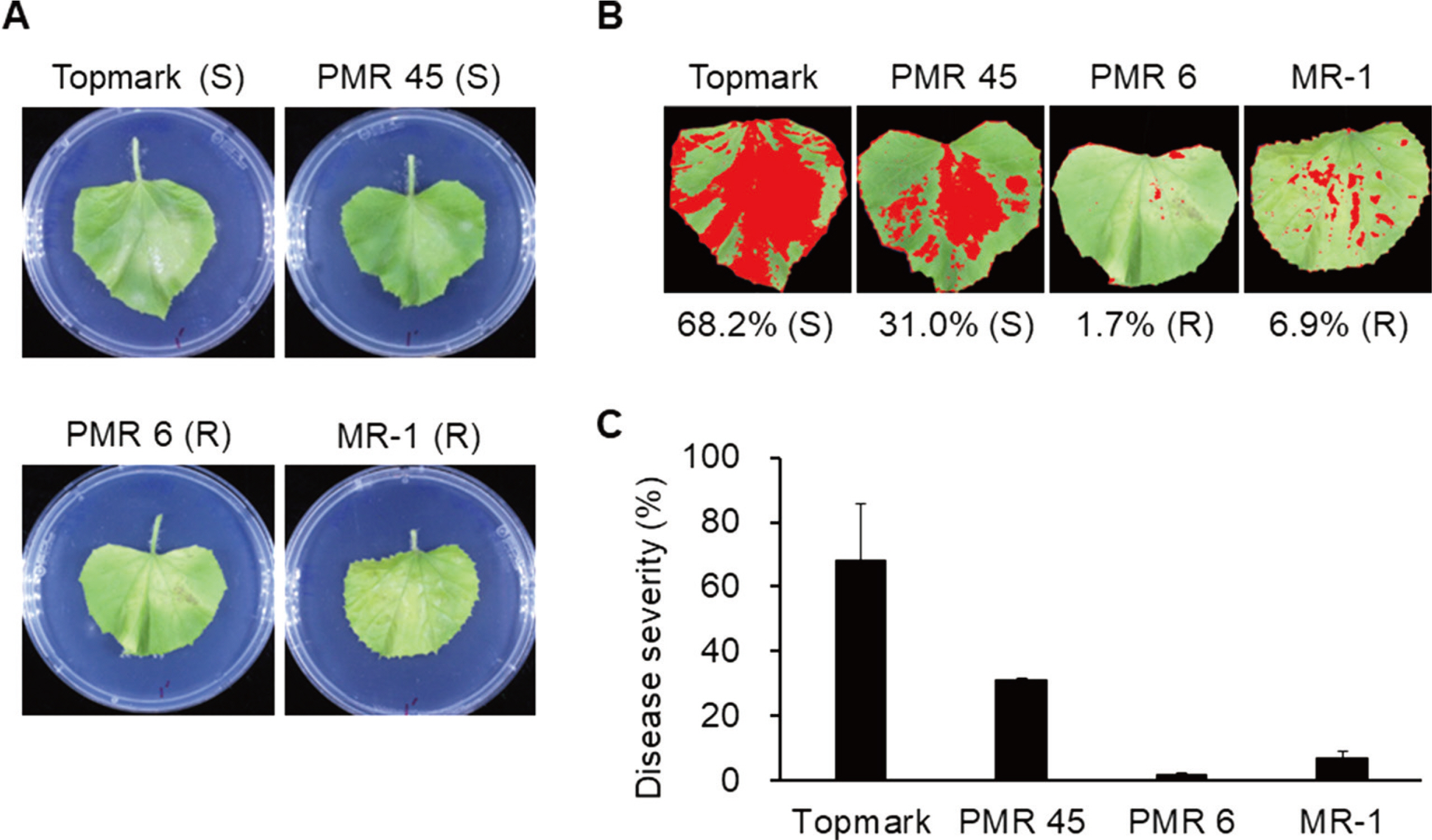
ļČäļ”¼ ĻĘĀņŻ╝ SE180328ņØś ņĀĆĒĢŁņä▒ Ļ▓ĆņĀĢ Ļ▓░Ļ│╝, ŌĆśTopmarkŌĆÖņŚÉņä£ 68.2% (S), ŌĆśPMR 45ŌĆÖņŚÉņä£ 31.0% (S), ŌĆśPMR 6ŌĆÖņŚÉņä£ 1.7% (R) ĻĘĖļ”¼Ļ│Ā ŌĆśMR-1ŌĆÖņŚÉņä£ 6.9% (R)ņØś ļ░£ļ│æ ņĀĢļÅäļź╝ ļéśĒāĆļé┤ņŚłļŗż(Fig. 4). ņÜ░ļ”¼ļŖö ļ│Ė Ļ▓ĆņĀĢ Ļ▓░Ļ│╝ļź╝ ĻĖ░ņĪ┤ņŚÉ ļ│┤Ļ│ĀļÉśņŚłļŹś ņŚ░ĻĄ¼ Ļ▓░Ļ│╝(Lee ļō▒, 2014; Zitter ļō▒, 1996)ņÖĆ ļ╣äĻĄÉĒĢśņŚ¼ SE180328ļź╝ race 2ļĪ£ Ļ▓░ļĪĀ ļé┤ļĀĖļŗż(Table 1). ņ¦ĆĻĖłĻ╣īņ¦Ć ĻĄŁļé┤ņØś P. xanthiiņØś Ļ▓ĮņÜ░ race 2ļĪ£ ļČäļźś ļ│┤Ļ│ĀļÉ£ ņśłĻ░Ć ņŚåņ¢┤ ņāłļĪ£ņÜ┤ raceĻ░Ć ņ£Āņ×ģ Ēś╣ņØĆ ļ│ĆņØ┤ĒĢśĻ│Ā ņ׳ņØīņØä ņŗ£ņé¼ĒĢśņśĆļŗż.
ņČöĻ░Ć Ļ▓ĆņĀĢņØä ĒåĄĒĢ£ Race 2FņØś ļ│┤Ļ│Ā
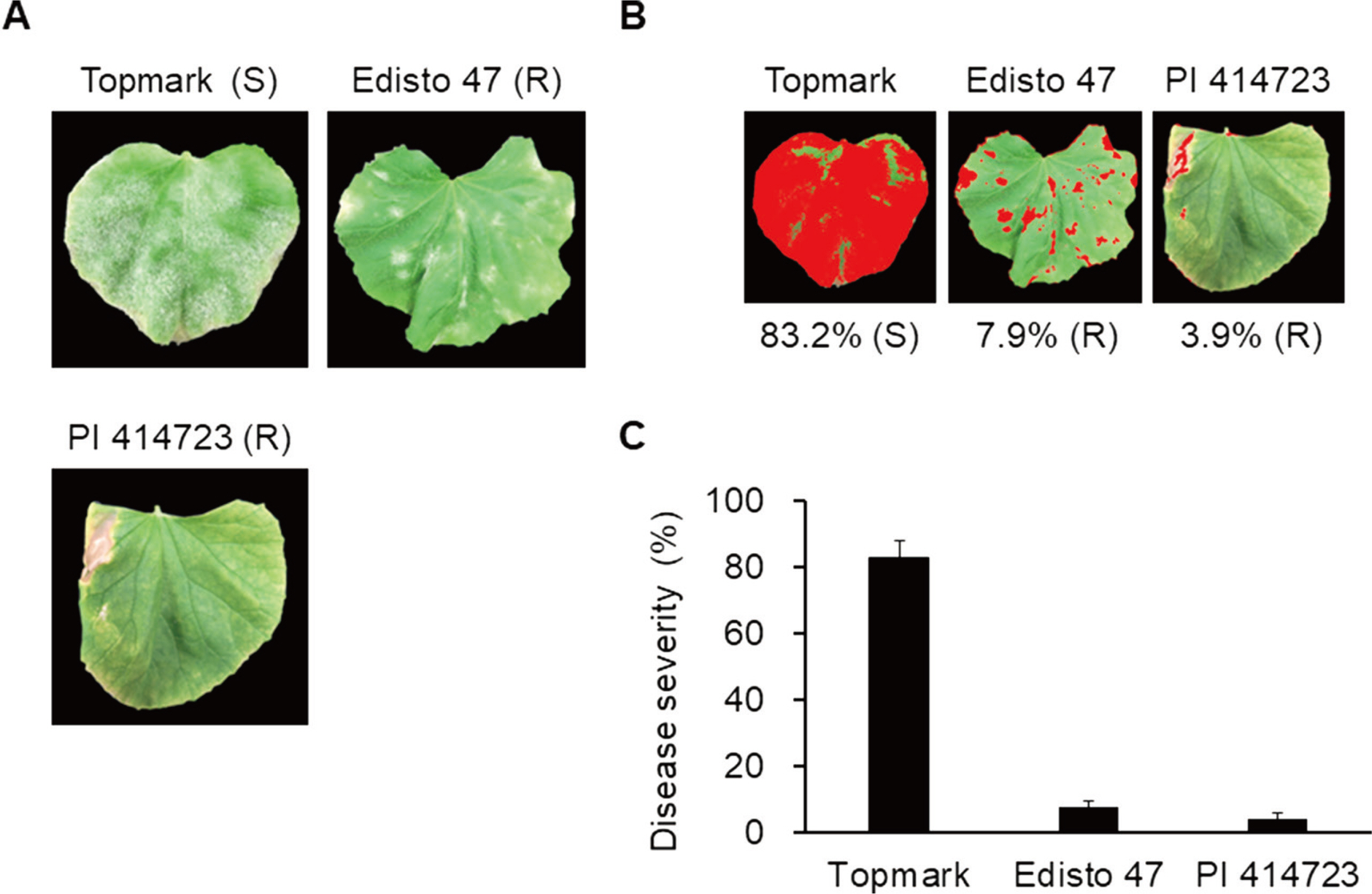
P. xanthii race 1Ļ│╝ race 2ņØś ļ│ĆņØ┤ņ▓┤(variants)ņŚÉ ļīĆĒĢ£ ņŚ░ĻĄ¼ļź╝ ļ│┤ļ®┤ race 1ņŚÉņä£ 8Ļ░£ņØś ļ│ĆņØ┤ņ▓┤ņÖĆ race 2ņŚÉņä£ 6Ļ░£ņØś ļ│ĆņØ┤ņ▓┤Ļ░Ć ļ│┤Ļ│ĀļÉśņ¢┤ ņ׳ļŗż(Cohen ļō▒, 2004; McCreight, 2006). Race 2ņØś ļ│ĆņØ┤ņ▓┤ņØĖ 2aņÖĆ 2bņØś Ļ▓ĮņÜ░ ņĢäņ¦ü ļ¬ģĒÖĢĒĢ£ ņŚ░ĻĄ¼Ļ░Ć ļÉśņ¦Ć ņĢŖņĢä ļŗżļźĖ ļ│ĆņØ┤ņ▓┤ņÖĆ ļÅÖņØ╝ĒĢĀ Ļ░ĆļŖźņä▒ņØ┤ ņĀ£ĻĖ░ļÉśĻ│Ā ņ׳ļŗż(Cohen ļō▒, 2000; McCreight, 2006). ņĄ£ĻĘ╝ņŚÉļŖö 10Ļ░£ņØś ļ®£ļĪĀ ĒīÉļ│ä ņŗØļ¼╝ņØä ņØ┤ņÜ®ĒĢśņŚ¼ race 2ļź╝ 2S (Salinas), 2US(United States), 2F (France), 2SC (South Carolina)ļĪ£ ļČäļźśĒĢśņśĆļŗż(Tetteh ļō▒, 2013). ļ»ĖĻĄŁņŚÉņä£ļŖö P. xanthii race 2USĻ░Ć ņłśļ░ĢņŚÉņä£ ņóĆ ļŹö ļ¦ÄņØĆ Ēö╝ĒĢ┤ļź╝ ņØ╝ņ£╝ĒéżļŖö Ļ▓āņ£╝ļĪ£ ļ│┤Ļ│ĀļÉśņŚłņ£╝ļ®░(Davis ļō▒, 2007; Tetteh ļō▒, 2013), ņżæĻĄŁņŚÉņä£ļŖö ļ®£ļĪĀņŚÉ ĒØ░Ļ░ĆļŻ©ļ│æņØä ņØ╝ņ£╝ĒéżļŖö ņŻ╝ņÜö P. xanthiiĻ░Ć race 2Fņ×äņØ┤ ļ░ØĒśĆņĀĖ(Wang ļō▒, 2006), ĻĄŁĻ░Ćļ│äļĪ£ ņ×æļ¬®ļ│äļĪ£ Ļ░üĻĖ░ ņāüņØ┤ĒĢ£ raceĻ░Ć ņÜ░ņĀÉĒĢśĻ│Ā ņ׳ļŖö Ļ▓āņ£╝ļĪ£ ļ│┤Ļ│ĀļÉśĻ│Ā ņ׳ļŗż. ņØ┤ņŚÉ ĻĖ░ņĪ┤ņØś ņŚ░ĻĄ¼ļōżņØä(Cosme ļō▒, 2012; McCreight, 2006; Tetteh ļō▒, 2013) ļö░ļØ╝ņä£ ņČöĻ░ĆņĀüņØĖ ļ®£ļĪĀ ĒīÉļ│ä ņŗØļ¼╝ņØĖ ŌĆśEdisto 47ŌĆÖņÖĆ ŌĆśPI 414723ŌĆÖņØä ņØ┤ņÜ®ĒĢśņŚ¼ SE180328Ļ░Ć race 2ņØś ļ│ĆņØ┤ņ▓┤ņØĖ 2S, 2US, 2F, 2SC ņżæņŚÉņä£ ņ¢┤ļ¢ż raceņØĖņ¦Ćļź╝ ņĪ░ņé¼ĒĢśņśĆļŗż(Fig. 5). TopmarkļŖö ņŚ¼ņĀäĒ׳ ļåÆņØĆ Ļ░Éņłśņä▒(83.2%)ņØä ļ│┤ņśĆņ£╝ļéś ŌĆśEdisto 47ŌĆÖņÖĆ ŌĆśPI 414723ŌĆÖņØĆ Ļ░üĻ░ü 7.9%ņÖĆ 3.9%ļĪ£ ļ¬©ļæÉ ļåÆņØĆ ņłśņżĆņØś ņĀĆĒĢŁņä▒ņØä ļéśĒāĆļāłļŗż. ņØ┤ļź╝ ļ░öĒāĢņ£╝ļĪ£ SE180328ļŖö P. xanthii race 2FļĪ£ ņĄ£ņóģ Ļ▓░ļĪĀĒĢśņśĆļŗż(Table 1).
ņÜö ņĢĮ
ĒØ░Ļ░ĆļŻ©ļ│æ(powdery mildew)ņØĆ ņśżņØ┤(Cucumis sativus)ļź╝ ĒżĒĢ©ĒĢśļŖö ļ░ĢĻ│╝(Cucurbitaceae) ņ×æļ¼╝ņŚÉņä£ ņĀä ņäĖĻ│äņĀüņ£╝ļĪ£ Ļ░Ćņן ņŗ¼Ļ░üĒĢ£ Ēö╝ĒĢ┤ļź╝ ņØ╝ņ£╝ĒéżļŖö ņŻ╝ņÜöĒĢ£ ļ│æ ņżæ ĒĢśļéśņØ┤ļŗż. ņśżņØ┤ ĒØ░Ļ░ĆļŻ©ļ│æņØś ņ¦ĆņåŹņĀüņØĖ ļ░£ļ│æĻ│╝ ļ░®ņĀ£ņØś ņ¢┤ļĀżņøĆ ļĢīļ¼ĖņŚÉ ĻĘĖ ņøÉņØĖ ļ│æņøÉĻĘĀņØä ļ░ØĒ׳Ļ│Ā raceļź╝ ĻĘ£ļ¬ģĒĢśļŖö Ļ▓āņØ┤ ĒĢäņłśņĀüņØ┤ļŗż. ĻĄŁļé┤ņŚÉņä£ ļ░£ļ│æĒĢśļŖö ņśżņØ┤ ĒØ░Ļ░ĆļŻ©ļ│æĻĘĀņØś ņĪ░ņé¼ļź╝ ņ£äĒĢśņŚ¼ Ļ▓ĮĻĖ░ļÅä Ļ┤æņŻ╝(Gwangju), Ļ▓ĮĻĖ░ļÅä ņØ┤ņ▓£(Icheon), ņĀäļØ╝ļČüļÅä Ļ╣ĆņĀ£(Gimje), ņä£ņÜĖĒŖ╣ļ│äņŗ£(Seoul), Ļ▓Įņāüļé©ļÅä ļ░Ćņ¢æ(Miryang) ņ¦ĆņŚŁņŚÉņä£ ĒØ░Ļ░ĆļŻ©ļ│æņØ┤ ļ░£ņāØĒĢ£ ņśżņØ┤ ņ×ÄņØä ņ▒äņ¦æĒĢśņśĆļŗż. ĒśĢĒā£ĒĢÖņĀü, ļČäņ×ÉņāØļ¼╝ĒĢÖņĀü ĒŖ╣ņä▒ ņĪ░ņé¼ Ļ▓░Ļ│╝ ļČäļ”¼ļÉ£ ņśżņØ┤ ĒØ░Ļ░ĆļŻ©ļ│æĻĘĀļōżņØĆ ņĀłļīĆ ĒÖ£ļ¼╝ĻĖ░ņāØĻĘĀņØĖ Podosphaera xanthiiļĪ£ ļÅÖņĀĢļÉśņŚłļŗż. P. xanthiiņØś race ĻĘ£ļ¬ģņŚÉ ņé¼ņÜ®ļÉśĻ│Ā ņ׳ļŖö ļ®£ļĪĀ(C. melo) ĒīÉļ│ä ĒÆłņóģņØä ņØ┤ņÜ®ĒĢśņŚ¼ ļ░Ćņ¢æ, ņØ┤ņ▓£ ņä£ņÜĖņŚÉņä£ ļČäļ”¼ļÉ£ MI180427, IC190611, SE180328ņØś ļĀłņØ┤ņŖżļź╝ Ļ░üĻ░ü ņĪ░ņé¼ĒĢśņśĆļŗż. ļČäļ”¼ĻĘĀņŻ╝ MI180427Ļ│╝ IC190611ļŖö ĻĄŁļé┤ņŚÉņä£ Ļ░Ćņן Ļ┤æļ▓öņ£äĒĢśĻ▓ī ļ░£ļ│æļÉśĻ│Ā ņ׳ļŖö Ļ▓āņ£╝ļĪ£ ļ│┤Ļ│ĀļÉ£ race 1ņ£╝ļĪ£ ĻĘ£ļ¬ģļÉśņŚłļŗż. ļ░śļ®┤, ļČäļ”¼ĻĘĀņŻ╝ SE180328ļŖö ĻĄŁļé┤ņŚÉņä£ ņĢäņ¦üĻ╣īņ¦Ć ļ│┤Ļ│ĀļÉ£ ņśłĻ░Ć ņŚåļŖö race 2ļĪ£ ĒīÉļ│äļÉśņŚłļŗż. SE180328ņØś ņĀĢĒÖĢĒĢ£ race ĒīÉļ│äņØä ņ£äĒĢ┤ņä£ ņČöĻ░ĆņĀüņØĖ ļ®£ļĪĀ ĒīÉļ│ä ĒÆłņóģņŚÉ ņĀæņóģĒĢ£ Ļ▓░Ļ│╝, ĒØ░Ļ░ĆļŻ©ļ│æ race 2 ņżæņŚÉņä£ ņżæĻĄŁ ļ▓ĀņØ┤ņ¦ĢņØś ņŻ╝ņÜö raceļĪ£ ņĢīļĀżņ¦ä race 2FļĪ£ ļÅÖņĀĢļÉśņŚłļŗż. ļö░ļØ╝ņä£ ņāłļĪ£ņÜ┤ race 2FņØś ĻĄŁļé┤ ļ░£ļ│æņØä ļ│┤Ļ│ĀĒĢśļŖö ļ│Ė ņŚ░ĻĄ¼ Ļ▓░Ļ│╝ļŖö Ē¢źĒøä ņśżņØ┤ ĒØ░Ļ░ĆļŻ©ļ│æ ņĀĆĒĢŁņä▒ ĒÆłņóģ ņ£Īņä▒ņŚÉ Ļ│ĀļĀż ļÉśņ¢┤ņĢ╝ ĒĢĀ ĻĖ░ņ┤łņ×ÉļŻīĻ░Ć ļÉĀ Ļ▓āņ£╝ļĪ£ ņāØĻ░üļÉ£ļŗż.


 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






