


ņä£ ļĪĀ
Ļ│ĀņČö(Capsicum spp.)ļŖö ņŚ┤ļīĆņä▒ ņŗØļ¼╝ļĪ£ ņÜ░ļ”¼ļéśļØ╝ņŚÉņä£ ņŻ╝ņÜö Ļ▓Į ņĀ£ņ×æļ¼╝ļĪ£ ņ×Éļ”¼ ņ×ĪĻ│Ā ņ׳Ļ│Ā, ņĪ░ļ»Ė ņ▒äņåīļźś ņżæ Ļ░Ćņן ļäōņØĆ ņ×¼ļ░░ļ®┤ņĀü ņØä ņ░©ņ¦ĆĒĢśĻ│Ā ņ׳ļŗż. ĻĘĖļ¤¼ļéś ņĄ£ĻĘ╝ ĻĖ░ņāüņØ┤ļ│ĆĻ│╝ ņ×¼ļ░░ ļåŹļ»╝ņØś Ļ│ĀļĀ╣ĒÖö ļō▒ņŚÉ ļö░ļØ╝ ņ×¼ļ░░ņØś ņ¢┤ļĀżņøĆņØä Ļ▓¬Ļ│Ā ņ׳ņØä ļ┐Éļ¦ī ņĢäļŗłļØ╝, ņÜ░ļ”¼ļéśļØ╝ņŚÉ ņä£ļŖö ņŚ░ņ×æ, ņåīĻĘ£ļ¬© ņ×¼ļ░░, ļģĖļÅÖ ņ¦æņĢĮ, ĻĖ┤ ņ×¼ļ░░ ņ×æĻĖ░ļĪ£ ņØĖĒĢ┤ ņŚ¼ļ¤¼ ļ│æ ĒĢ┤ Ēö╝ĒĢ┤Ļ░Ć Ēü░ ņ×æļ¼╝ņØ┤ĻĖ░ļÅä ĒĢśļŗż(Park ļō▒, 2012). ņÜ░ļ”¼ļéśļØ╝ņŚÉ ļ│┤Ļ│Ā ļÉ£ Ļ│ĀņČöņŚÉ ļ░£ņāØĒĢśļŖö ļ│æĒĢ┤ļŖö 35ņóģņØ┤ļ®░, ĻĘĖ ņżæ ņŚŁļ│æĻ│╝ ĒāäņĀĆļ│æ, ļ¬© ņ×ÉņØ┤Ēü¼ļ│æ ļō▒ ņĢĮ 10ņóģņØś ļ│æņØ┤ ļ¦żļģä Ļ│ĀņČöņŚÉ Ēü░ Ēö╝ĒĢ┤ļź╝ ņŻ╝ļŖö Ļ▓āņ£╝ ļĪ£ ļ│┤Ļ│ĀļÉśņŚłĻ│Ā, Ļ│ĀņČöĒāäņĀĆļ│æĻ│╝ ņŚŁļ│æņØ┤ Ļ░Ćņן ņŗ¼Ļ░üĒĢ£ Ēö╝ĒĢ┤ļź╝ ņØ╝ņ£╝ ĒéżĻ│Ā ņ׳ļŗż(Seo ļō▒, 2011; The Korean Society of Plant Pathology, 2009). Ļ│ĀņČöĒāäņĀĆļ│æĻĘĀ ņżæ ņłśļ¤ēĻ│╝ ĒÆłņ¦łņŚÉ ņ¦üņĀæņĀüņØĖ ņśüĒ¢źņØä ļ»Ėņ╣śļŖö ĻĘĀņØĆ C. acutatumĻ│╝ C. gloeosporioidesļĪ£ ņĢīļĀżņĀĖ ņ׳ņ£╝ ļ®░ ņĄ£ĻĘ╝ņŚÉļŖö C. acutatumņØ┤ ņŻ╝ļĪ£ ļ░£ņāØĒĢ£ļŗżĻ│Ā ļ│┤Ļ│ĀļÉśņŚłļŗż(Kim ļō▒, 2008). ņŻ╝ļĪ£ ļ»ĖņłÖĒĢ£ Ļ│╝ņŗżļĪ£ļČĆĒä░ ņłśĒĢÖ ĒøäĻ╣īņ¦Ć ļ░£ņāØĒĢśļ®░ ņłśņ╣© ņāüņ£╝ļĪ£ ņØīĒæ╣ ļōżņ¢┤Ļ░ä ņøÉĒśĢ ļ░śņĀÉņ£╝ļĪ£ ļéśĒāĆļéśĻ│Ā ļ│æļ░ś ņ£ŚļČĆļČäņŚÉ ļŗ┤ ĒÖ®ņāē ļé┤ņ¦Ć ĒĢæĒü¼ņāēņØ┤ļéś ņśżļĀīņ¦ĆņāēņØś Ēżņ×É ļŹ®ņ¢┤ļ”¼ ņ”ØņāüņØä ļ│┤ņØ┤ļ®░ ņŗØļ¼╝ņ▓┤ ņĀä ņāØņ£ĪĻĖ░ņŚÉ Ļ▒Ėņ│É Ļ│ĀņČö Ļ│╝ņŗżņŚÉņä£ Ļ░ĢĒĢ£ ļ│æņøÉņä▒ņØ┤ ļ│┤Ļ│ĀļÉ£ ļ░ö ņ׳ļŗż(Han ļō▒, 2009).
ĒāäņĀĆļ│æ ļ░®ņĀ£ļź╝ ņ£äĒĢ┤ ņé¼ņÜ®ĒĢśļŖö ĒÖöĒĢÖļåŹņĢĮņØĆ dithianon, carbendazium, chlorothalonill, azoxystrobin, mancozeb ļō▒ņØ┤ ņ׳ ņ¦Ćļ¦ī, ņāØĒā£Ļ│ä ĒīīĻ┤┤ņÖĆ Ļ░ÖņØĆ ļ¼ĖņĀ£ņĀÉĻ│╝ C. acutatumņØś ņĢĮņĀ£ ļé┤ņä▒ ļō▒ņ£╝ļĪ£ ņØĖĒĢśņŚ¼ ļ░®ņĀ£ņŚÉ ņ¢┤ļĀżņøĆņØ┤ ņ׳ļŗż(Kwak ļō▒, 2012). ņØ┤ņŚÉ ņĀĆ ļÅģņä▒ņØ┤ļ®┤ņä£ ĒÖśĻ▓ĮņŚÉ ĒĢ┤Ļ░Ć ņŚåļŖö ņāØļ¼╝ņĀü ļ░®ņĀ£ņŚÉ ņ׳ņ¢┤ņä£ ĒŖ╣Ē׳ ļ│æņøÉ ĻĘĀņŚÉ ĒĢŁĻĘĀļĀźņØä ļ│┤ņØ┤ļŖö ļ»ĖņāØļ¼╝ņØä ņØ┤ņÜ®ĒĢ£ ņāØļ¼╝ĒĢÖņĀü ļ░®ņĀ£Ļ░Ć Ļ┤Ćņŗ¼ ņØä ļüīĻ│Ā ņ׳ņ£╝ļ®░, ĒåĀņ¢æņŚÉņä£ Bacillus sp., Psedomonas sp., Strepto- myces sp. ļō▒Ļ│╝ Ļ░ÖņØĆ ļ»ĖņāØļ¼╝ņØä ļČäļ”¼ĒĢśņŚ¼ ĒĢŁņāØļ¼╝ņ¦łĻ│╝ ļ»ĖņāØļ¼╝ņĀ£ ņØś Ļ░£ļ░£ņØä ņ£äĒĢ£ ņŚ░ĻĄ¼ļōżņØ┤ ņ¦ĆņåŹņĀüņ£╝ļĪ£ ņØ┤ļŻ©ņ¢┤ņ¦ĆĻ│Ā ņ׳ļŗż(KimĻ│╝ Yun, 2011; Lee, 2010; Paik ļō▒, 1996; Park ļō▒, 2006). ĻĘĖ ņżæņŚÉņä£ļÅä ņŗØļ¼╝ņāØņ£Īņ┤ēņ¦ä ĻĘ╝ĻČīņäĖĻĘĀņ£╝ļĪ£ ļé┤ņāØĒżņ×É(endospores)ļź╝ ĒśĢņä▒ĒĢś ņŚ¼ ņןĻĖ░Ļ░ä ļ│┤ņĪ┤ņØ┤ Ļ░ĆļŖźĒĢ£ BacillusņóģņŚÉ ļīĆĒĢ£ Ļ░£ļ░£ņØ┤ ņ”ØĻ░ĆĒĢśĻ│Ā ņ׳ ļŗż(Borriss, 2011; Qiao ļō▒, 2014).
ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö Ļ│ĀņČöĒāäņĀĆļ│æņØä ņØ╝ņ£╝ĒéżļŖö C. acutatumņŚÉ ļīĆĒĢ£ ņāØļ¼╝ĒĢÖņĀü ļ░®ņĀ£ĒÜ©Ļ│╝Ļ░Ć ņÜ░ņłśĒĢ£ BacillusņóģņØä ņäĀļ░£ĒĢśĻ│Ā ĒāäņĀĆļ│æņØś ļ░£ļ│æ ņ¢ĄņĀ£ ĒÜ©Ļ│╝ ļ░Å ņŗØļ¼╝ņāØņןņ┤ēņ¦äļŖźņØä Ļ▓ĆņĀĢĒĢśņśĆņ£╝ļ®░, ņŗżņĀ£ ļåŹĻ░Ć ņŚÉņä£ ĻĖ░ņĪ┤ ĒÖöĒĢÖļåŹņĢĮĻ│╝ņØś Ēś╝ņÜ®ņØ┤ Ļ░ĆļŖźĒĢ£ ņ¦Ć ņŚ¼ļČĆņŚÉ ļīĆĒĢśņŚ¼ Ļ▓ĆĒåĀ ĒĢśĻ│Ā ņØ┤ļź╝ ļ│┤Ļ│ĀĒĢśĻ│Āņ×É ĒĢ£ļŗż.
ņ×¼ļŻī ļ░Å ļ░®ļ▓Ģ
ĻĘĀņŻ╝ņäĀļ░£.
ņØĖņé╝ņ×¼ļ░░ņ¦Ć ĒåĀņ¢æņ£╝ļĪ£ļČĆĒä░ ņ£ĀņÜ®ļ»ĖņāØļ¼╝ņØä ļČäļ”¼ĒĢśņśĆļŗż. ņØĖņé╝ ĻĘ╝ĻČīņØś ĒåĀņ¢æņØä ņ▒äņĘ©ĒĢśņŚ¼ ļ®ĖĻĘĀņłśņŚÉ ĒśäĒāüĒĢśņŚ¼ ĒؼņäØ Ēøä brain heart infusion (BHI) ļ░░ņ¦ĆņŚÉ ļÅäļ¦ÉĒĢśĻ│Ā ļŗ©ņØ╝ ņĮ£ļĪ£ļŗłņØś ļ»ĖņāØļ¼╝ļōżņØä ļČäļ”¼ĒĢśņśĆļŗż(Park ļō▒, 2013). ĻĖĖĒĢŁļĀźņØ┤ ņÜ░ņłśĒĢ£ ļ»ĖņāØļ¼╝ ņäĀļ░£ņØä ņ£äĒĢ┤, ņØĖņé╝ļ┐īļ”¼ņŹ®ņØīļ│æņøÉĻĘĀ Cylindrocarpon destructans Ēżņ×É ĒśäĒāØņĢĪņØä 105 conidia/mlņØś ļåŹļÅäļĪ£ ņĀ£ņĪ░ĒĢśņŚ¼ potato dextrose agar (PDA) ļ░░ņ¦ĆņŚÉ ļÅäļ¦ÉĒĢ£ Ēøä ņāüĻĖ░ņŚÉņä£ ļČäļ”¼ĒĢ£ ļŗ©ņØ╝ ņĮ£ļĪ£ļŗłņØś ļ»ĖņāØļ¼╝ļōżņØä Ļ░üĻ░ü ņé┤ĻĘĀļÉ£ ņØ┤ņæżņŗ£Ļ░£ļĪ£ ļ¼╗ĒśĆ ņĀæņóģĒĢśĻ│Ā 21┬░CņŚÉņä£ 5ņØ╝Ļ░ä ļ░░ņ¢æĒĢśņŚ¼ inhibition zoneņØ┤ ĒśĢņä▒ļÉ£ ņĮ£ļĪ£ļŗłļź╝ ņäĀļ░£ĒĢśņśĆļŗż.
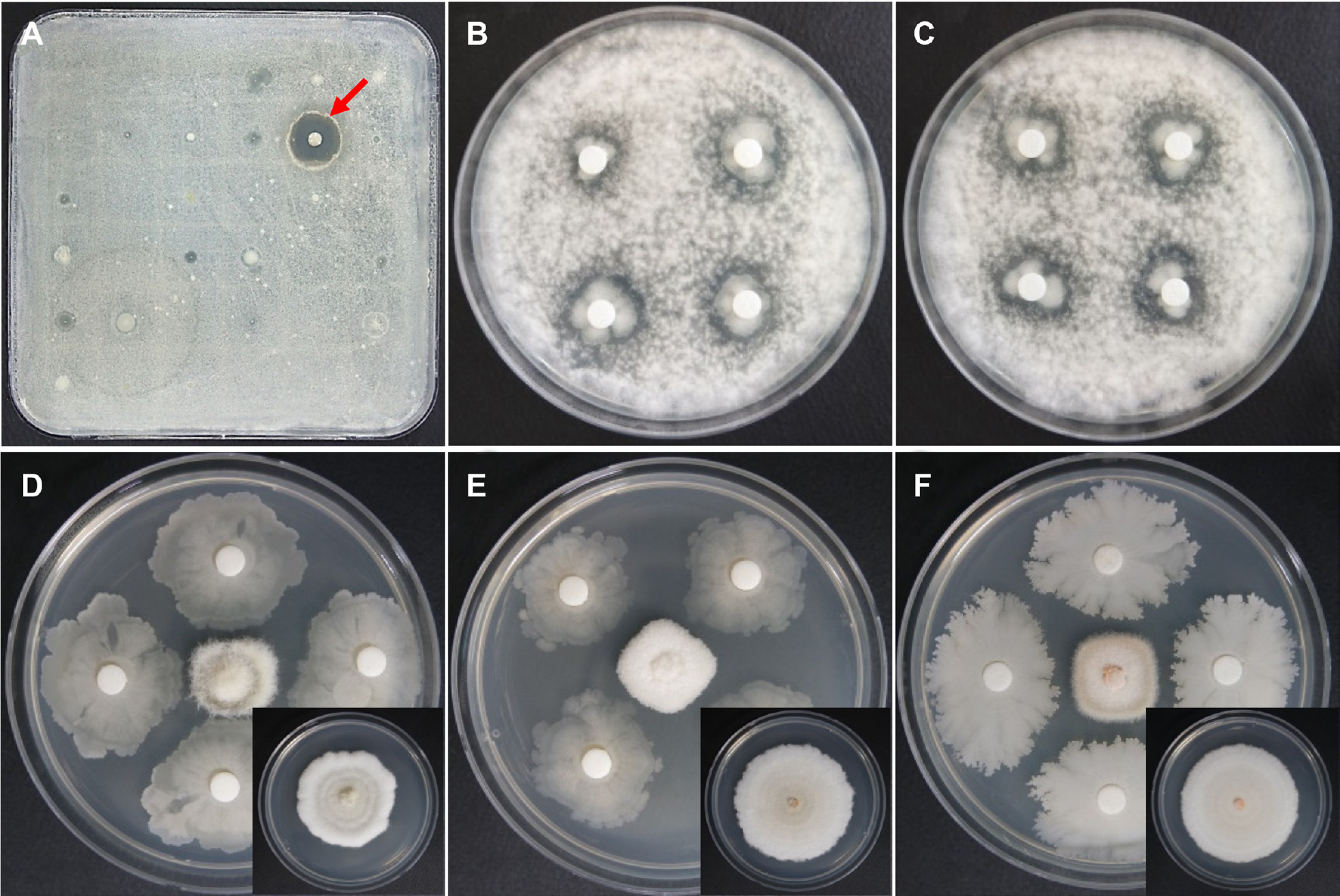
Ļ│ĀņČö ĒāäņĀĆļ│æĻĘĀņØĆ ĒĢ£ĻĄŁļåŹņŚģļ»ĖņāØļ¼╝ņ×ÉņøÉņä╝Ēä░ļĪ£ļČĆĒä░(Korean Agricultural Culture Collection, KACC) ļČäņ¢æļ░øņĢä ņé¼ņÜ®ĒĢśņśĆņ£╝ļ®░(Collectotrichum acutatum KACC40847, Collectotrichum acutatum KACC42509, Collectotrichum acutatum KACC42403 ļ░Å Collectotrichum gloeosporioides KACC42690), ņØ┤ļōż ĻĘĀņŻ╝ņØś ļ░░ņ¢æļ░░ņ¦ĆņŚÉ 6-mm cork borerļĪ£ Ļ│ĀņČöĒāäņĀĆļ│æ ļĖöļ¤ŁņØä ņלļØ╝ ņāłļĪ£ņÜ┤ PDA ļ░░ņ¦Ć ņżæņĢÖņŚÉ ņ╣śņāüĒĢśĻ▒░ļéś, Ēżņ×ÉĒśäĒāüņĢĪ (105 conidia/ml) 100 ┬ĄlņØä ņĀ£ņĪ░ĒĢśņŚ¼ PDA ļ░░ņ¦ĆņŚÉ Ļ░üĻ░ü ļÅäļ¦ÉĒĢ£ Ēøä, 6-mm paper discļź╝ ņ╣śņāüĒĢśĻ│Ā ĻĘĖ ņ£äņŚÉ ņäĀļ░£ļÉ£ ĻĘĀņŻ╝ ļ░░ņ¢æņĢĪ 10 ┬ĄlņØä ņĀæņóģĒĢśņŚ¼ inhibition zone ĒśĢņä▒ ņŚ¼ļČĆļź╝ Ļ┤Ćņ░░ĒĢśņśĆļŗż.
16S rDNA ņŚ╝ĻĖ░ņä£ņŚ┤ ļČäņäØ.
YGB36ņØś 16S ribosomal RNA ņ£ĀņĀäņ×É ņä£ņŚ┤ņØś PCR ņ”ØĒÅŁņØä ņ£äĒĢ┤, 27F (5ŌĆ▓-AGAGTTTGATCMTG- GCTCAG-3ŌĆ▓)ņÖĆ 1492R (5ŌĆ▓-GGYTACCTTGTTACG ACTT-3ŌĆ▓) ĒöäļØ╝ņØ┤ļ©Ėļź╝ ņé¼ņÜ®ĒĢśņśĆņ£╝ļ®░, ņ”ØĒÅŁļÉ£ PCR ņé░ļ¼╝ņØś sequencingņØä ņłśĒ¢ē ĒĢ£ Ēøä NCBI (National Center for Biotechnology Information) ņØś BLASTļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ļČäņäØĒĢśņśĆļŗż. Phylogenetic ļČäņäØņŚÉļŖö MEGA-XĻ░Ć ņé¼ņÜ®ļÉśņŚłļŗż(Kumar ļō▒, 2016; Weisburg ļō▒, 1991).
Biolog, Vitek systemņØä ņØ┤ņÜ®ĒĢ£ ļÅÖņĀĢ.
ņäĀļ░£ļÉ£ ĻĘĀņŻ╝ņØś ĒāäņåīņøÉ ņØ┤ņÜ®ļÅäļź╝ ņØ┤ņÜ®ĒĢ£ ļÅÖņĀĢņØä ņ£äĒĢ┤ Biolog GEN III microplate system (Biolog, Hayward, CA, USA)ņØä ņé¼ņÜ®ĒĢśņśĆļŗż. ņ¦Ćņ╣©ņŚÉ ļö░ļØ╝, ĻĘĀņŻ╝ ņĮ£ļĪ£ļŗłļź╝ fluid A (Biolog)ņŚÉ ĒśäĒāü Ēøä Ļ░ü 100 ┬Ąlļź╝ GEN III microplateņØś ļ¬©ļōĀ wellņŚÉ ņĀæņóģĒĢśņŚ¼, 28┬░CņŚÉņä£ 42ņŗ£Ļ░ä ļ░░ņ¢æĒĢśņśĆņ£╝ļ®░, Biolog MicroStation automated readerļź╝ ņé¼ņÜ®ĒĢśņŚ¼ ņĖĪņĀĢ Ēøä MicroLog 3 software (Biolog)ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ļČäņäØĒĢśņśĆļŗż. ĒĢ£ĒÄĖ, ĒĢ┤ļŗ╣ ĻĘĀņŻ╝ņØś ļÅÖņĀĢņØä ņ£äĒĢ┤ Vitek systemņØ┤ ņé¼ņÜ®ļÉśņŚłļŗż. Vitek 2 Compact system (bioM├®rieux, France)ņŚÉņä£ BCLņ╣┤ļō£ļź╝ ņé¼ņÜ®ĒĢśņŚ¼ ņĀ£ņĪ░ņé¼ņØś ņ¦Ćņ╣©ņŚÉ ļö░ļØ╝ ņ¦äĒ¢ēļÉśņŚłņ£╝ļ®░, 28┬░CņŚÉņä£ 14ņŗ£Ļ░ä ļ░░ņ¢æ Ēøä VITEK 2 Compact software version 07.01ņ£╝ļĪ£ ļČäņäØĒĢśņśĆļŗż.
ļ░░ņ¢æĒŖ╣ņä▒.
ļ░░ņ¢æ ĒŖ╣ņä▒ņØä ņĪ░ņé¼ĒĢśĻĖ░ ņ£äĒĢśņŚ¼, YGB36ņØä BHI ņĢĪņ▓┤ ļ░░ņ¦ĆņŚÉ ņĀæņóģĒĢ£ Ēøä 28┬░CņŚÉņä£ 7ņØ╝Ļ░ä ļ░░ņ¢æĒĢśļ®┤ņä£, UV 600 nmņŚÉņä£ņØś ODĻ░ÆņØä ļ¦żņØ╝ ņĖĪņĀĢĒĢśņśĆņ£╝ļ®░, ņĄ£ņ┤ł 24ņŗ£Ļ░äĻ╣īņ¦ĆļŖö 6ņŗ£Ļ░ä Ļ░äĻ▓®ņ£╝ļĪ£ ņĖĪņĀĢĒĢśņśĆļŗż. ļÅÖņŗ£ņŚÉ ņāØĻĘĀņłś ņĖĪņĀĢņØä ņ£äĒĢ┤, ODĻ░Æ ņĖĪņĀĢļĢī ļ¦łļŗż ļ░░ņ¢æņĢĪņØś ņØ╝ļČĆļź╝ ņĘ©ĒĢśņŚ¼ BHI Ļ│Āņ▓┤ļ░░ņ¦ĆņŚÉ 10-fold dilution platingĒĢśņŚ¼ ļ░░ņ¢æĒĢ£ Ēøä ļ░£ņāØĒĢ£ ņĮ£ļĪ£ļŗł ņłśļź╝ ņĖĪņĀĢĒĢśņśĆļŗż.
ĒÜ©ņåīĒÖ£ņä▒ Ļ▓ĆņĀĢ.
Cellulase ņāØņé░ ņŚ¼ļČĆļź╝ ņĢīņĢäļ│┤ĻĖ░ ņ£äĒĢ┤ CMC agar ļ░░ņ¦Ć(carboxyl methyl cellulose 5 g, MgSO4 2 g, CaCl2 0.5 g, KCl 1 g, FeSO4 0.001 g, agar 20 g)ļź╝ ņØ┤ņÜ®ĒĢśņśĆļŗż(Sazci ļō▒, 1986). ļ®ĖĻĘĀļÉ£ 6-mm filter paperļź╝ ļ░░ņ¦Ć ņ£ä 3Ļ││ņŚÉ ņś¼ļ”░ Ēøä 2Ļ││ ņŚÉļ¦ī YGB36 ļ░░ņ¢æņĢĪņØä 10 ┬Ąlņö® ņĀæņóģĒĢ£ Ēøä 28┬░CņŚÉņä£ 5ņØ╝Ļ░ä ļ░░ņ¢æĒĢśņśĆļŗż. 0.1% Congo red ņÜ®ņĢĪņ£╝ļĪ£ ļ░░ņ¦Ćļź╝ ņŚ╝ņāēĒĢ£ Ēøä inhibition zone ĒśĢņä▒ņØä Ļ┤Ćņ░░ĒĢśņśĆļŗż. ļ│Ė ņŗżĒŚśņØĆ 3ļ░śļ│Ąņ£╝ļĪ£ ņ¦äĒ¢ēĒĢśņśĆļŗż.
Protease ĒÖ£ņä▒ņØĆ gelatinņØä ņ▓©Ļ░ĆĒĢśņŚ¼ ļ¦īļōĀ ĒÅēĒīÉļ░░ņ¦Ć(gelatin 5 g, beef extract 3 g, proteose peptone 5 g, agar 15 g)ņŚÉ YGB36 ļ░░ņ¢æņĢĪņØä ņĀæņóģĒĢśĻ│Ā 5ņØ╝Ļ░ä ļ░░ņ¢æĒĢ£ Ēøä, 1% tannic acid ņÜ®ņĢĪņ£╝ļĪ£ ņŚ╝ņāēĒĢśņŚ¼ inhibition zone ĒśĢņä▒ņØä Ļ┤Ćņ░░ĒĢśņśĆļŗż. ļ│Ė ņŗżĒŚśņØĆ 3ļ░śļ│Ąņ£╝ļĪ£ ņ¦äĒ¢ēĒĢśņśĆļŗż.
Amylase ĒÖ£ņä▒ņØĆ Ļ░ĆņÜ®ņä▒ ņĀäļČä(soluble starch)ņØä ņ▓©Ļ░ĆĒĢśņŚ¼ ļ¦īļōĀ ĒÅēĒīÉļ░░ņ¦Ć(beef extract 3 g, proteose peptone 5 g, soluble starch 2 g, agar 15 g)ņŚÉ YGB36 ļ░░ņ¢æņĢĪņØä ņĀæņóģĒĢśĻ│Ā 5ņØ╝Ļ░ä ļ░░ņ¢æ ĒĢ£ Ēøä, GramŌĆÖs iodine ņÜ®ņĢĪ(3.3 g iodine crystal, 2.8 g potassium iodine, 1,000 ml distilled water)ņ£╝ļĪ£ ņŚ╝ņāēĒĢśņŚ¼ inhibition zone ĒśĢņä▒ņØä Ļ┤Ćņ░░ĒĢśņśĆļŗż. ļ│Ė ņŗżĒŚśņØĆ 3ļ░śļ│Ąņ£╝ļĪ£ ņ¦äĒ¢ēĒĢśņśĆļŗż.
Siderophore ņāØņé░ ņŚ¼ļČĆļŖö ļ│ĆĒśĢļÉ£ Chrome azurol S (CAS) agar assayļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņĖĪņĀĢĒĢśņśĆļŗż(Han ļō▒, 2015). CAS (Sigma, St. Louis, MO, USA) ņŚ╝ļŻī ņÜ®ņĢĪņØĆ ņ”Øļźśņłś 50 mlņŚÉ CAS 60.5 mg ņØä ļģ╣ņØ┤Ļ│Ā, 72.9 mgņØś hexadecyltrimethylammonium bro- mideļź╝ ņ”Øļźśņłś 40 mlņŚÉ ļģ╣ņØ┤Ļ│Ā, Luria-Bertani agar 35 gņØä ņ”Øļźśņłś 900 mlņŚÉ ļģ╣ņØĖ Ēøä ņäĖ Ļ░Ćņ¦Ć ņÜ®ņĢĪņØä Ēś╝ĒĢ®ĒĢśĻ│Ā Ļ│ĀņĢĢ ļ®ĖĻĘĀĒĢśņśĆļŗż. HCl ņÜ®ņĢĪ(10 mM) 10 mlņŚÉ 1 mM FeCl3ŌĆó6H2Oļź╝ ļģ╣ņŚ¼ ļö░ļĪ£ Ļ│ĀņĢĢļ®ĖĻĘĀĒĢśņśĆļŗż. Ļ│ĀņĢĢ ļ®ĖĻĘĀļÉ£ ļæÉ ņÜ®ņĢĪņØä 50┬░CļĪ£ ņŗØĒ×ī Ēøä ņä×ņ¢┤ņŻ╝Ļ│Ā Petri dishņŚÉ ļČäņŻ╝ĒĢśņŚ¼ CAS ĒÅēĒīÉļ░░ņ¦Ćļź╝ ļ¦īļōżņŚłļŗż. ņØ┤Ēøä ņ£äņÖĆ Ļ░ÖņØĆ ļ░®ļ▓Ģņ£╝ļĪ£ ņäĖĻĘĀņØä ņĀæņóģĒĢśĻ│Ā 28┬░CņŚÉņä£ 5ņØ╝Ļ░ä ļ░░ņ¢æĒĢ£ Ēøä, orange halo zone ĒśĢņä▒ņØä Ļ┤Ćņ░░ĒĢśņśĆļŗż. ļ│Ė ņŗżĒŚśņØĆ 3ļ░śļ│Ąņ£╝ļĪ£ ņ¦äĒ¢ēĒĢśņśĆļŗż.
ņØĖņé░ ļČäĒĢ┤ļŖź(phosphate solubilization)ņØĆ tricalcium phosphateĻ░Ć ĒżĒĢ©ļÉ£ ļ░░ņ¦Ć(glucose 10 g, tricalcium phosphate 5 g, MgCl2ŌĆó6H2O 5 g, MgSO4ŌĆó7H2O 0.25 g, KCl 0.2 g, (NH4)2SO4 0.1 g, agar 20 g)ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼, ņāüĻĖ░ņÖĆ Ļ░ÖņØĆ ļ░®ļ▓Ģņ£╝ļĪ£ ņäĖĻĘĀ ļ░░ņ¢æņĢĪņØä ņĀæņóģĒĢśĻ│Ā 15ņØ╝Ļ░ä ļ░░ņ¢æĒĢ£ Ēøä halo zone ĒśĢņä▒ņØä Ļ┤Ćņ░░ĒĢśņśĆļŗż. ļ│Ė ņŗżĒŚśņØĆ 3ļ░śļ│Ąņ£╝ļĪ£ ņ¦äĒ¢ēĒĢśņśĆļŗż.
In vitro Ēżņ×É ļ░£ņĢä ņ¢ĄņĀ£ ĒÜ©Ļ│╝.
YGB36 ļ░░ņ¢æņĢĪ ļ░Å ļ░░ņ¢æņŚ¼ņĢĪņØä ņØ┤ņÜ®ĒĢśņŚ¼ C. acutatum KACC42403ņØś Ēżņ×Éļ░£ņĢä ņ¢ĄņĀ£ ĒÜ©Ļ│╝ļź╝ ņĪ░ņé¼ĒĢśņśĆļŗż. C. acutatumņØĆ PDA ļ░░ņ¦ĆņŚÉņä£ 5ņØ╝Ļ░ä ļ░░ņ¢æĒĢśņŚ¼ Ēżņ×É ĒśäĒāüņĢĪņØä 105 conidia/ml ļåŹļÅäļĪ£ ņżĆļ╣äĒĢśņśĆĻ│Ā, YGB36ņØĆ BHI ņĢĪņ▓┤ļ░░ņ¦ĆņŚÉņä£ 3ņØ╝Ļ░ä ļ░░ņ¢æĒĢ£ ļ░░ņ¢æņĢĪ ļ░Å ņØ┤ļź╝ MF-Millipore membrane filter (pore size, 0.22 ┬Ąm)ļĪ£ ņŚ¼Ļ│╝ĒĢśņŚ¼ ļ░░ņ¢æņŚ¼ņĢĪņ£╝ļĪ£ ņżĆļ╣äĒĢśņśĆļŗż. ņŖ¼ļØ╝ņØ┤ļō£ ĻĖĆļØ╝ņŖż ņ£äņŚÉ Ēżņ×ÉĒśäĒāüņĢĪ 10 ┬ĄlņØä ļČäņŻ╝ĒĢśĻ│Ā ņØ┤ņŚÉ ļ░░ņ¢æņĢĪ ļśÉļŖö ļ░░ņ¢æņŚ¼ņĢĪ 10 ┬Ąlļź╝ Ļ░üĻ░ü ļČäņŻ╝ĒĢśņśĆļŗż. ņŖĄļÅäļź╝ ņ£Āņ¦ĆĒĢśļ®░ 25┬░CņŚÉņä£ ļ░░ņ¢æĒĢśļ®┤ņä£ 8ņŗ£Ļ░ä Ļ░äĻ▓®ņ£╝ļĪ£ 48ņŗ£Ļ░ä ļÅÖņĢł ļ░£ņĢä(germination) ņ£Āļ¼┤ņÖĆ ļČĆņ░®ĻĖ░(appressorium) ĒśĢņä▒ņØä ņĪ░ņé¼ĒĢśņśĆļŗż. Ēżņ×ÉņØś ļ░£ņĢäļŖö ļ░£ņĢäĻ┤Ć(germ tube)ņØś ĻĖĖņØ┤Ļ░Ć Ēżņ×É Ēü¼ĻĖ░ņØś 1/2 ņØ┤ņāü ļÉśņŚłņØä ļĢīļĪ£ ņĀĢņØśĒĢśņśĆņ£╝ļ®░, ļīĆņĪ░ĻĄ¼ļĪ£ļŖö ļ®ĖĻĘĀņłśļź╝ ņé¼ņÜ®ĒĢśņŚ¼ ņ£äņÖĆ Ļ░ÖņØĆ ļ░®ļ▓Ģņ£╝ļĪ£ ņĪ░ņé¼ĒĢśņśĆļŗż. Ēżņ×É ļ░£ņĢäņ£©ņØĆ (1-ļ░£ ņĢäņ£©/ļīĆņĪ░ĻĄ¼ļ░£ņĢäņ£©)├Ś100ņ£╝ļĪ£ Ļ│äņé░ĒĢśņśĆņ£╝ļ®░, ļ¬©ļōĀ ņŗżĒŚśņØĆ 3ļ░śļ│Ąņ£╝ļĪ£ ņŗżņŗ£ĒĢśņśĆļŗż.
Ļ│ĀņČöĒāäņĀĆļ│æ ļ░£ņāØņ¢ĄņĀ£.
ņŗ£ņżæņŚÉņä£ ĒīÉļ¦żļÉśļŖö ĒÆŗĻ│ĀņČöļź╝ 70% ethanolĻ│╝ 2% NaOCl ņØä ņØ┤ņÜ®ĒĢ£ Ēæ£ļ®┤ ņé┤ĻĘĀ Ēøä ņé¼ņÜ®ĒĢśņśĆļŗż. ļ®ĖĻĘĀļÉ£ ĒĢĆņØä ņØ┤ņÜ®ĒĢśņŚ¼ Ļ│ĀņČö Ļ│╝ņŗż Ēæ£ļ®┤ 6Ļ││ņŚÉ ņāüņ▓śļź╝ ļéĖ ļÆż C. acutatum KACC42403 Ēżņ×ÉĒśäĒāüņĢĪ(105 conidia/ml) 10 ┬Ąlļź╝ ņĀæņóģĒĢ£ Ēøä ņŗżņś©ņŚÉņä£ Ļ▒┤ņĪ░ĒĢśņśĆļŗż. YGB36ņØĆ BHI ņĢĪņ▓┤ļ░░ņ¦ĆņŚÉ 3ņØ╝Ļ░ä ļ░░ņ¢æ ĒĢ£ Ēøä 20ļ░░ļĪ£ ĒؼņäØĒĢśņŚ¼ ļČäļ¼┤ĒĢśņśĆĻ│Ā ļīĆņĪ░ĻĄ¼ļŖö BHI ņĢĪņ▓┤ļ░░ņ¦Ćļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ļČäļ¼┤ĒĢśņśĆļŗż Ļ│ĀņČö Ļ│╝ņŗżņØĆ ņŖĄņŗżņ▓śļ”¼ĒĢśņŚ¼ 10ņØ╝Ļ░ä 25┬░C ļĪ£ ļ░░ņ¢æĒĢśļ®┤ņä£ ļ│æ ļ░£ņāØņØä ļ╣äĻĄÉĒĢśņśĆļŗż. ļ¬©ļōĀ ņŗ£ĒŚśņØĆ 3ļ░śļ│Ąņ£╝ļĪ£ ņ¦äĒ¢ēĒĢśņśĆļŗż. ļ░£ļ│æļÅä(%)ļŖö ļ│æļ░ś Ēü¼ĻĖ░ņŚÉ ļö░ļØ╝ ļ░£ļ│æņ¦Ćņłśļź╝ 0, 0% ļ│æņ¦ĢņŚåņØī; 1, <2 mm; 2, 2-4 mm; 3, 5-7 mm; 4, 8-10 mm; 5, 10mm ņ┤łĻ│╝ļĪ£ ļéśļłäņ¢┤ ĒÅēĻ░ĆĒĢśņśĆĻ│Ā, ļ░£ļ│æļÅäļŖö {Ōłæ(ļ░£ņāØņ¦Ćņłś├Śļ░£ļ│æņłś)/5(ņĄ£Ļ│Āļ░£ļ│æņ¦Ćņłś)├Ś6(ņĪ░ņé¼ņłś)}├Ś100ļĪ£ ņé░ņČ£ĒĢśņśĆļŗż. ļ░®ņĀ£Ļ░ĆļŖö (ļīĆņĪ░ĻĄ¼ļ░£ļ│æļÅä-ņ▓śļ”¼ĻĄ¼ļ░£ļ│æļÅä/ļīĆņĪ░ĻĄ¼ļ░£ļ│æ)├Ś100ņ£╝ļĪ£ ņé░ņČ£ ĒĢśņśĆļŗż.
Ļ│ĀņČö ņóģņ×Éļ░£ņĢä ļ░Å ņāØņ£Īņ┤ēņ¦äĒÜ©Ļ│╝.
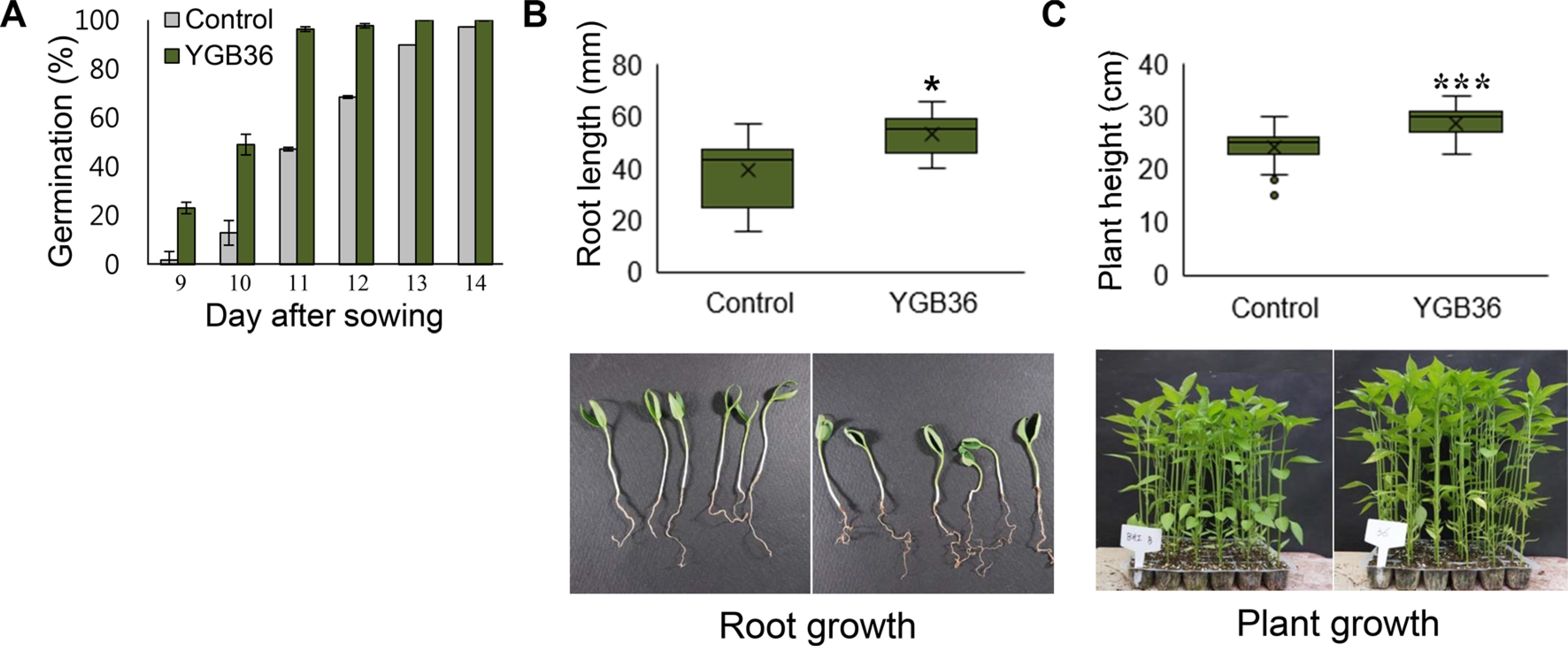
YGB36ņØś Ļ│ĀņČö ņāØņ£Ī ņ┤ēņ¦ä ĒÜ©Ļ│╝ ņŗżĒŚśņØĆ ņĢłļÅÖļīĆĒĢÖĻĄÉ ļČĆņåŹļåŹņן ņŗżĒŚśņŗżņŖĄĒż ņ£Āļ”¼ņś©ņŗżņŚÉņä£ ņ¦äĒ¢ēĒĢśņśĆļŗż. 36ĻĄ¼ ĒżĒŖĖ(27.5 cm├Ś27.5 cm├Ś3.5 cm)ņŚÉ ņāüĒåĀļź╝ ņ▒äņÜ┤ ļÆż Ļ│ĀņČöņóģņ×É(Dogyacheongcheong, Syngenta Korea, Seoul, Korea)ļź╝ ĒīīņóģĒĢśņśĆĻ│Ā, YGB36ņØĆ BHIļ░░ņ¦ĆņŚÉņä£ 3ņØ╝Ļ░ä ļ░░ņ¢æ Ēøä ļ®ĖĻĘĀņłśļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ 10ļ░░ ĒؼņäØĒĢśņŚ¼ 7ņØ╝ Ļ░äĻ▓®ņ£╝ļĪ£ Ļ┤ĆņŻ╝ĒĢśņśĆļŗż. ļīĆņĪ░ĻĄ¼ļĪ£ļŖö BHI ņĢĪņ▓┤ļ░░ņ¦Ćļź╝ ņé¼ņÜ®ĒĢśņśĆļŗż. ļ░£ņĢäņ£© Ļ┤Ćņ░░ņØĆ Ēīīņóģ Ēøä 9ņØ╝ļČĆĒä░ 14ņØ╝Ļ╣īņ¦Ć, ļ┐īļ”¼ ņāØņ£Ī ņĖĪņĀĢņØĆ 15ņØ╝ ņ░©ņŚÉ, ņ¦ĆņāüļČĆ ĻĖĖņØ┤ ņĖĪņĀĢņØĆ 58ņØ╝ ņ░©ņŚÉ ņ¦äĒ¢ēĒĢśņśĆļŗż. ļ¬©ļōĀ ņ▓śļ”¼ĻĄ¼ļŖö 36ĻĄ¼ ĒżĒŖĖņØś 3ļ░śļ│Ąņ£╝ļĪ£ ņŗżņŗ£ĒĢśņśĆļŗż.
ĒÖöĒĢÖļåŹņĢĮņŚÉ ļīĆĒĢ£ ņĀĆĒĢŁņä▒ Ļ▓ĆņĀĢ.
ņŗżĒŚśņØä ņ£äĒĢ┤ Ēśäņ×¼ ņ▓ŁņåĪņ¦ĆņŚŁ Ļ│ĀņČöņ×¼ļ░░ ļåŹĻ░ĆņŚÉņä£ ņĄ£ĻĘ╝ 3ļģäĻ░ä ņé¼ņÜ®ļÉ£ ņé┤ĻĘĀņĀ£ 21ņóģĻ│╝ ņé┤ņČ®ņĀ£ 14ņóģņØä ņØ┤ņÜ®ĒĢśņśĆļŗż(Table 1). 3ņØ╝Ļ░ä ņĢĪņ▓┤ ļ░░ņ¢æĒĢ£ YGB36ņØś ļ░░ ņ¢æņĢĪ 100 ┬Ąlļź╝ BHI ļ░░ņ¦ĆņŚÉ ļÅäļ¦ÉĒĢ£ Ēøä ņĀüņĀłĒĢ£ Ļ░äĻ▓®ņ£╝ļĪ£ 4Ļ░£ņØś 6 mm paper discļź╝ ņś¼ļ”¼Ļ│Ā, ĒÖöĒĢÖļåŹņĢĮņØĆ ņĀ£ņĪ░ņé¼ņØś ņ¦Ćņ╣©ņŚÉņä£ ņĀ£ņŗ£ĒĢśļŖö ļåŹļÅä(├Ś1)ļź╝ ņżæņŗ¼ņ£╝ļĪ£ ļ░śļ¤ē(├Ś0.5) ļ░Å ļ░░ļ¤ē(├Ś2)ņ£╝ļĪ£ ņĀ£ņĪ░ĒĢśņŚ¼ Ļ░ü 10 ┬Ąlņö® ļČäņŻ╝ĒĢśņśĆļŗż. 3ņØ╝Ļ░ä ļ░░ņ¢æ Ēøä inhibition zoneņØä Ļ┤Ćņ░░ĒĢśņśĆļŗż. ļ¬©ļōĀ ņŗżĒŚśņØĆ 3ļ░śļ│Ąņ£╝ļĪ£ ņ¦äĒ¢ēĒĢśņśĆļŗż.
ĒåĄĻ│äļČäņäØ.
Ļ░ü ņ▓śļ”¼ļ│ä ĒÅēĻĘĀ Ļ░ä ņ░©ņØ┤ņŚÉ ļīĆĒĢ£ ĒåĄĻ│äļŖö ļČäņé░ļČäņäØ ņ£╝ļĪ£ ņ¦äĒ¢ēĒĢśņśĆĻ│Ā, Ļ░ü ņ▓śļ”¼ Ļ░äņØś ņ░©ņØ┤ļŖö ņĄ£ņåīņ£ĀņØśņ░© Ļ▓ĆņĀĢ(least significant difference [LSD] test)ņ£╝ļĪ£ ļŗżņżæ ļ╣äĻĄÉĒĢśņśĆņ£╝ļ®░, RĒåĄĻ│ä ņåīĒöäĒŖĖņø©ņ¢┤(R Foundation for Statistical Computing, Vienna, Austria)ļź╝ ņé¼ņÜ®ņśĆĒĢśļŗż.
Ļ▓░Ļ│╝ ļ░Å Ļ│Āņ░░
ĻĘĀņŻ╝.
ņØĖņé╝ĒżņןņØś ĒåĀņ¢æņ£╝ļĪ£ļČĆĒä░ ņØĖņé╝ņŹ®ņØīļ│æĻĘĀņØĖ C. destructansņŚÉ ļīĆĒĢśņŚ¼ Ļ░ĢĒĢ£ ĒĢŁĻĘĀļĀźņØä ļ│┤ņØ┤ļŖö ĻĘĀņŻ╝ļź╝ ņäĀļ░£ĒĢśņśĆ Ļ│Ā(Fig 1A), ņØ┤ļź╝ YGB36ņ£╝ļĪ£ ļ¬ģļ¬ģĒĢśņśĆļŗż. YGB36 ĻĘĀņŻ╝ļŖö ņØ┤ņÖĖ ņŚÉļÅä ĒāäņĀĆļ│æĻĘĀņØĖ Colletotrichum acutatum 3ņóģ(KACC40847, KACC42403, KACC42509) ļ░Å Colletotrichum gloeosporioides 1 ņóģ(KACC42690)ņŚÉ ļīĆĒĢśņŚ¼ ĒĢŁĻĘĀĒÖ£ņä▒ņØä ļéśĒāĆļé┤ņŚłļŗż(Fig. 1B-F).
ļÅÖņĀĢ.
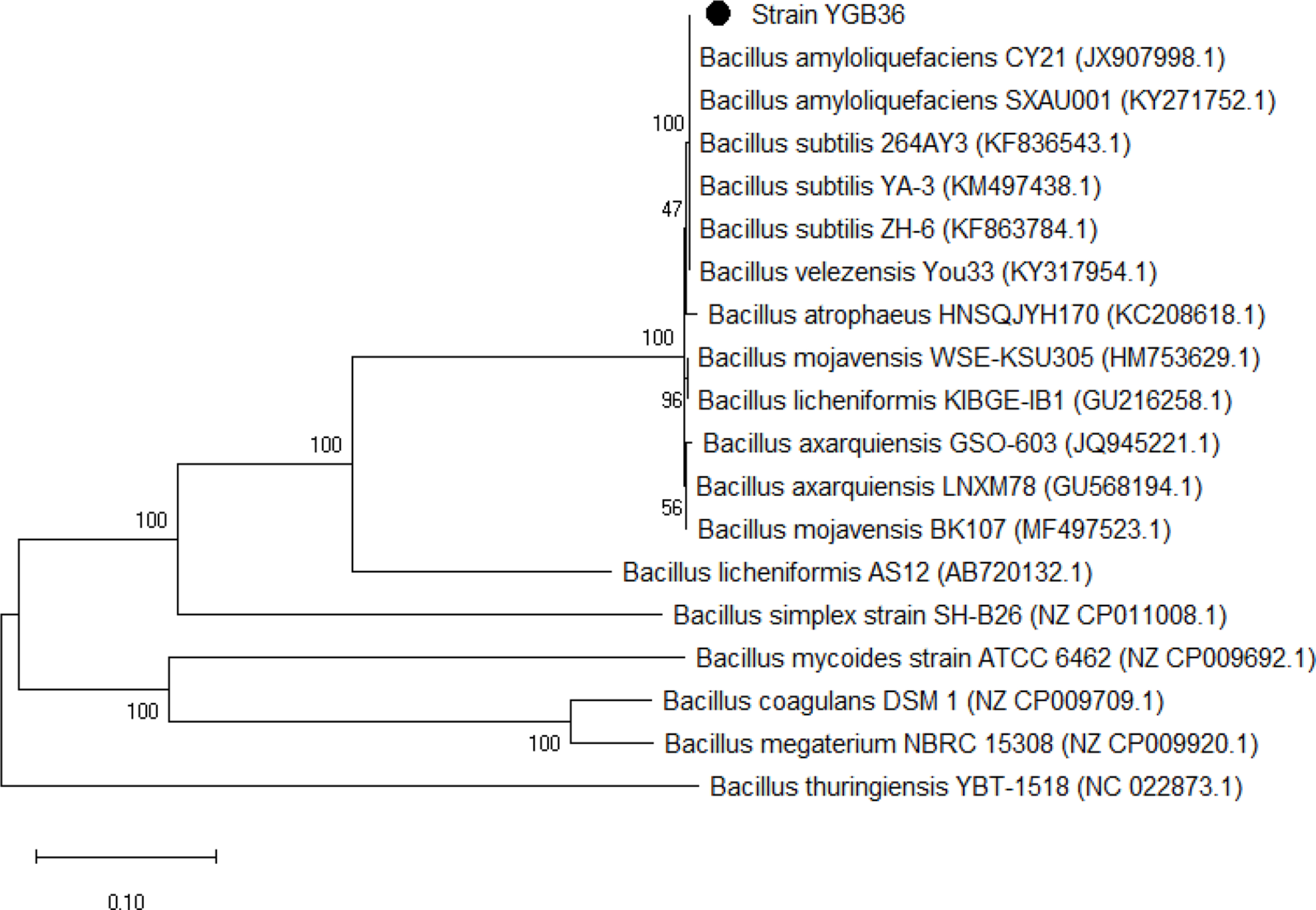
YGB36ĻĘĀņŻ╝ņØś Ļ▓īļåłDNAļź╝ ņČöņČ£ĒĢśņŚ¼ 16S ribosomal RNA ņ£ĀņĀäņ×Éļź╝ PCRļĪ£ ņ”ØĒÅŁĒĢśņŚ¼ ņĢĮ 1.2 kbņØś DNAņä£ņŚ┤ņØä ņ¢╗ņŚłņ£╝ļ®░, ņØ┤ņØś ņŚ╝ĻĖ░ņä£ņŚ┤ļČäņäØņØä ĒåĄĒĢ┤ Ļ│äĒåĄļČäļźśļź╝ ņ¦äĒ¢ēĒĢśņśĆļŗż(Fig. 2). ļČäņäØļÉ£ phylogenetic treeņŚÉņä£ ĒĢ┤ļŗ╣ ĻĘĀņŻ╝ļŖö Bacillus subtilis, Bacillus amyloliquefaciens ļ░Å Bacillus velezensisņÖĆ ļÅÖņØ╝ĒĢ£ ĻĘĖļŻ╣ņŚÉ ĒżĒĢ©ļÉśņŚłĻ│Ā, ņØ┤ļōżĻ│╝ļŖö ļ¬©ļæÉ 96.83%ņØś ņāüļÅÖņä▒ņØä ļéśĒāĆļé┤ņŚłļŗż. Biolog ļČäņäØņŚÉņä£, YGB36 ĻĘĀņŻ╝ļŖö Bacillus subtilisļĪ£ ļÅÖņĀĢļÉśņŚł ņ£╝ļ®░ ņØ┤ ļĢī probabilityļŖö 86.1%ņśĆĻ│Ā, Vitek 2 system ļČäņäØņŚÉņä£ļŖö, B. subtilis/B. amyloliquefaciens/B. atrophaeusļĪ£ ļÅÖņĀĢļÉśņŚłņ£╝ļ®░, probabilityļŖö 93%ņśĆļŗż. ĻĄ¼ņ▓┤ņĀüņ£╝ļĪ£, YGB36 ĻĘĀņŻ╝ļŖö GEN III microplateņāüņŚÉ ņ׳ļŖö substrates ņżæņŚÉņä£ 3-methyl glucose, D-serine, glucuronamide, ╬▒-keto-glutaric acid, ļ░Å D-malic acid ļō▒ņŚÉ ļīĆĒĢ┤ņä£ļŖö ļ░śņØæņØ┤ ļéśĒāĆļéśņ¦Ć ņĢŖņØĆ ļ░śļ®┤ņŚÉ myo-inositolĻ│╝ formic acid ļō▒ņŚÉ ļīĆĒĢ┤ņä£ļŖö ļ░śņØæņä▒ņØ┤ ļéśĒāĆļé¼ļŗż(Table 2). ņØ┤ļ¤¼ ĒĢ£ ļ░śņØæņä▒ņØĆ Bacillus amyloliquefaciensņÖĆļŖö ĻĄ¼ļ│äļÉśļŖö Bacillus subtilisņØś ĒŖ╣ņ¦ĢņØ┤ļŗż(VITEK 2 Compact software version 07.01). ĒĢ£ĒÄĖ, Vitek 2 system ļÅÖņĀĢņŚÉņä£ ĒĢ©Ļ╗ś ļČäļźśļÉ£ Bacillus velezensisļŖö Bacillus amyloliquefaciensņÖĆ ņāüĒśĖ ļ¦żņÜ░ ņ£Āņé¼ĒĢ┤ņä£, ļ¦ÄņØĆ Bacillus velezensisĻ░Ć ņØ┤ņĀäņŚÉļŖö Bacillus amyloliquefaciensļĪ£ ļČäļźśļÉśņŚłļŗżĻ░Ć ņĄ£ĻĘ╝ņŚÉņĢ╝ Bacillus velezensisļĪ£ ņ×¼ļČäļźśļÉśņ¢┤ ļ│┤Ļ│ĀļÉśĻ│Ā ņ׳ļŗż(Dunlap ļō▒, 2016; Wang ļō▒, 2008). ņóģĒĢ®ĒĢśļ®┤, 16S rRNA ņŚ╝ĻĖ░ņä£ņŚ┤ļČäņäØ, Biolog ļ░Å VitekļČäņäØņŚÉņä£ Ļ│ĄĒåĄņĀüņ£╝ļĪ£ B. subtilisĻ░Ć ļÅÖņĀĢļÉśņŚłĻ│Ā, ĒŖ╣ Ē׳ Biolog ļČäņäØņŚÉņä£ B. subtilis ĒŖ╣ņØ┤ņĀüņØĖ ĒŖ╣ņ¦ĢņØä ĒÖĢņØĖĒĢśņśĆņ£╝ļ»ĆļĪ£ YGB36 ĻĘĀņŻ╝ļŖö Bacillus subtilisļĪ£ ņĄ£ņóģ ļÅÖņĀĢļÉśņŚłļŗż.
ņāØņןĻ│ĪņäĀ ļ░Å ĒÜ©ņåīĒÖ£ņä▒.
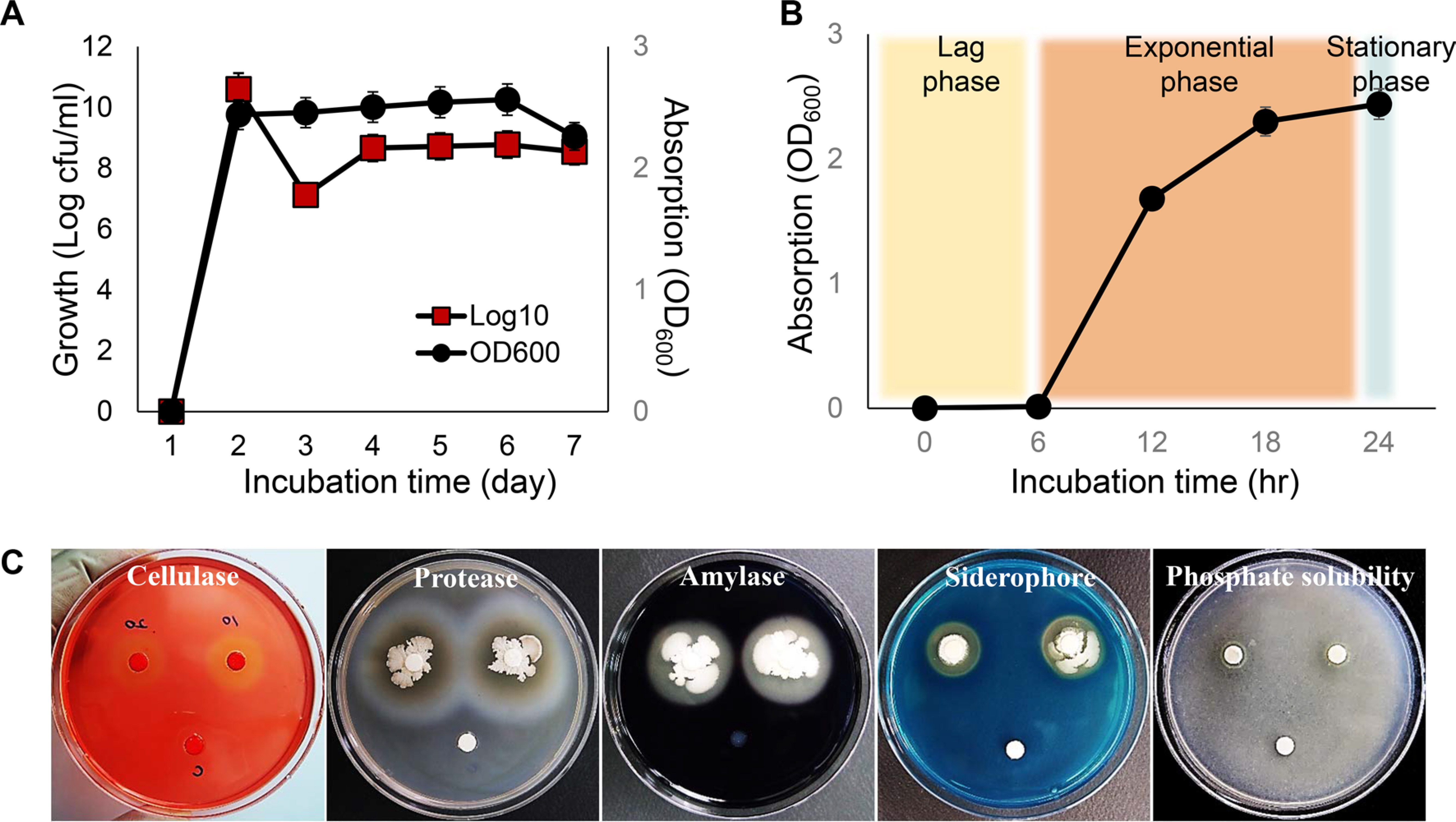
ņĢĪņ▓┤ļ░░ņ¦ĆņŚÉņä£ YGB36ņØä 7ņØ╝Ļ░ä ļ░░ņ¢æ ĒĢśļ®┤ņä£ ņāśĒöīļ¦üĒĢ£ Ēøä ļÅÖņØ╝ ņāśĒöīņŚÉ ļīĆĒĢśņŚ¼ ODĻ░Æ ņĖĪņĀĢ ļ░Å ņāØĻĘĀņłś Ļ│äņé░ņØä ĒåĄĒĢ┤ ņäĖĻĘĀņłśņØś ņ”ØņŗØņŚ¼ļČĆļź╝ ĒÖĢņØĖĒĢśĻ│Ā ĻĘĖļלĒöäļź╝ ņ¢╗ņŚłļŗż. ļæÉ ļ░®ļ▓Ģ ļ¬©ļæÉņŚÉņä£ ļ╣äņŖĘĒĢ£ Ļ▓░Ļ│╝ņØä ņ¢╗ņŚłņ£╝ļ®░, YGB36ņØä ņĀæņóģĒĢ£ņ¦Ć 24ņŗ£Ļ░ä ļ¦īņŚÉ stationary phaseņŚÉ ņ¦äņ×ģĒĢ£ Ļ▓āņØä ĒÖĢņØĖĒĢśņśĆĻ│Ā(Fig. 3A). YGB36ņØä ņĀæņóģĒĢ£ņ¦Ć ņĀüņ¢┤ļÅä 6ņŗ£Ļ░äĻ╣īņ¦ĆļŖö lag phaseĻ░Ć ņ¦ĆņåŹļÉ£ ĒøäņŚÉ exponential stageņŚÉ ņ¦äņ×ģĒĢśņśĆļŗż(Fig. 3B).
YGB36ņØś ĒĢŁĻĘĀĒÖ£ņä▒Ļ│╝ Ļ┤ĆļĀ©ļÉ£ ļŗżņ¢æĒĢ£ ĒÜ©ņåīĒÖ£ņä▒ņØä Ļ▓ĆņĀĢĒĢśņśĆļŗż(Fig. 3C). Cellulase ņāØņé░ ņŚ¼ļČĆ ņŗżĒŚśņØä ņ£äĒĢ┤ carboxymethyl celluloseļź╝ ņØ┤ņÜ®ĒĢśņśĆĻ│Ā, proteaseņŚÉ ļīĆĒĢ┤ņä£ļŖö gellatinņØä ņØ┤ņÜ® ĒĢśņśĆņ£╝ļ®░, amylase ņāØņé░ņØĆ soluble starchļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņĖĪņĀĢĒĢśņśĆļŗż. Ļ▓░Ļ│╝ņĀüņ£╝ļĪ£ YGB36ņØĆ cellulase, protease ļ░Å amylase ĒÖ£ņä▒ņØ┤ ļ¦żņÜ░ Ļ░ĢĒĢ£ Ļ▓āņ£╝ļĪ£ ļéśĒāĆļé¼ļŗż. Bacillus ņóģņŚÉņä£ ņØ┤ļ¤¼ĒĢ£ ĒÜ©ņåī ĒÖ£ņä▒ņØĆ ņØ┤ļ»Ė ļ│┤Ļ│ĀļÉśņ¢┤ ņ׳ņ£╝ļ®░, ļ│Ė ĻĘĀņŻ╝ņŚÉņä£ļÅä Ļ░ĢĒĢ£ ĒÖ£ņä▒ņØ┤ ĒÖĢņØĖļÉśņŚłļŗż(Bhaskar ļō▒, 2007; Nakamura ļō▒, 1987; Thippeswamy ļō▒, 2014).
ļŗżņØīņ£╝ļĪ£, ņŗØļ¼╝ņāØņןņ┤ēņ¦äņäĖĻĘĀņ£╝ļĪ£ņä£ņØś YGB36ĻĘĀņŻ╝ņØś ĒŖ╣ņä▒ņØä ņĢīņĢäļ│┤ĻĖ░ ņ£äĒĢśņŚ¼ siderophore ņāØņé░ ļ░Å ļé£ņÜ®ņä▒ņØĖņé░ņŚÉ ļīĆĒĢ£ ļČäĒĢ┤ļĀźņØä ņŗżĒŚśĒĢśņśĆļŗż. YGB36ņØĆ CAS agar ļ░░ņ¦ĆņŚÉņä£ halo zoneņØä ĒśĢņä▒ĒĢ©ņ£╝ļĪ£ņŹ© ļ╣äĻĄÉņĀü ļåÆņØĆ siderophoreļź╝ ņāØņé░ĒĢśĻ│Ā ņ׳ņŚłņ£╝ļ®░, tricalcium phosphateĻ░Ć ĒżĒĢ©ļÉ£ ļ░░ņ¦ĆņŚÉņä£ ļŗżņåīĻ░äņØś haloļź╝ ļ│┤ņŚ¼ ļČłņÜ®ņä▒ ņØĖņé░ņŚÉ ļīĆĒĢ£ ļČäĒĢ┤ļŖźļĀźņØ┤ ņ׳ņØīņØä ĒÖĢņØĖĒĢĀ ņłś ņ׳ņŚłļŗż. Plant growth-promoting rhizobacteria (PGPR)ņØĆ ņŗØļ¼╝ ĻĘ╝ĻČīņŚÉ ņĪ┤ņ×¼ĒĢśļ®┤ņä£ ņ▓Ā(Fe3+) ņä▒ļČä Ļ▓░ĒĢ®ļ¼╝ņ¦łņØĖ siderophoreļź╝ ņāØņé░ĒĢś ņŚ¼ ņ▓Ā ĒØĪņłśņ£©ņØä ļåÆņ×äņ£╝ļĪ£ņŹ© ĒåĀņ¢æņŚÉ ņ׳ļŹś ņ£ĀĒĢ┤ ļ»ĖņāØļ¼╝Ļ│╝ Ļ▓Įņ¤üņĀü ĻĖĖĒĢŁņ×æņÜ®ņØä ĒåĄĒĢ┤ ņśüĒ¢źņØä ņżä ņłś ņ׳ļŗż(BaggņÖĆ Neilands, 1987; Kloepper ļō▒, 1980). SiderophoreļŖö ņŗØļ¼╝ņØś ņ£ĀļÅäņĀäņŗĀņĀĆĒĢŁņä▒ (induced systemic resistance)ņŚÉļÅä Ļ┤ĆņŚ¼ĒĢśņŚ¼ ņŗØļ¼╝ņØś ļ│æņĀĆĒĢŁņä▒ņØä ļåÆņØ┤ļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż(Annapurna ļō▒, 2013; Beneduziļō▒, 2012). ļśÉĒĢ£ ļé£ņÜ®ņä▒ ņØĖņé░ņŚ╝ņØä ļČäĒĢ┤ĒĢśņŚ¼ ņŗØļ¼╝ņØ┤ ņ¦üņĀæ ņé¼ņÜ® ĒĢĀ ņłś ņ׳ļŖö ĒśĢĒā£ņØś ņśüņ¢æņøÉņ£╝ļĪ£ Ļ│ĄĻĖēĒĢ┤ņżīņ£╝ļĪ£ņŹ© ņŗØļ¼╝ņØś ņāØņןņ┤ēņ¦ä ĒÜ©Ļ│╝ļź╝ ļ│╝ ņłśļÅä ņ׳ļŗż(Rodr─▒guezņÖĆ Fraga, 1999). ņØ┤ņÖĆ Ļ░ÖņØ┤ YGB36ņØĆ ĒĢŁĻĘĀĒÖ£ņä▒Ļ│╝ ļŹöļČłņ¢┤ ņŗØļ¼╝ņāØņןņŚÉļÅä ļÅäņøĆņØä ņżä ņłś ņ׳ļŖö ņ£ĀņÜ®ĒĢ£ PGPRļĪ£ ņŚ¼Ļ▓©ņ¦äļŗż.
In vitro Ēżņ×Éļ░£ņĢäņ¢ĄņĀ£ĒÜ©Ļ│╝ Ļ▓ĆņĀĢ.
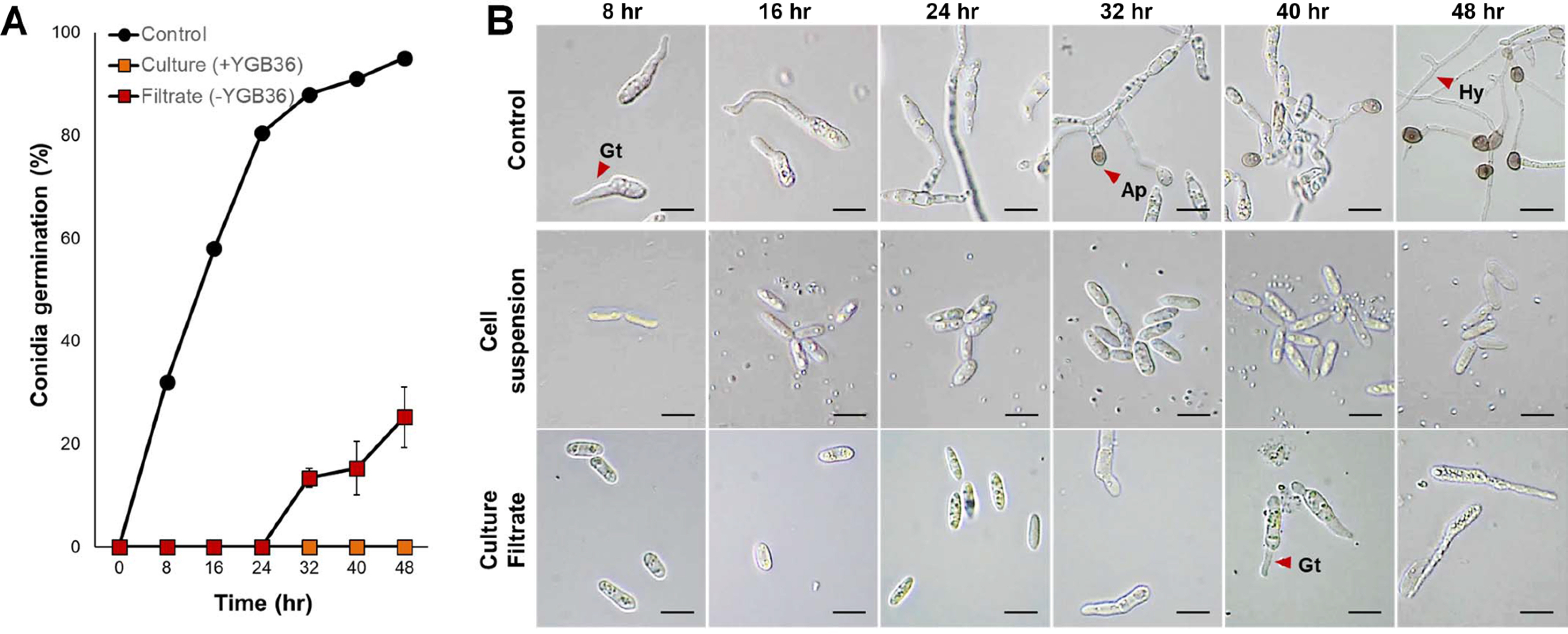
Ļ│ĀņČö ĒāäņĀĆļ│æĻĘĀņŚÉ ļīĆĒĢ£ YGB36ņØś ĒĢŁĻĘĀĒÖ£ņä▒ņØä ņĢīņĢäļ│┤ĻĖ░ ņ£äĒĢśņŚ¼ Ēżņ×Éļ░£ņĢäņ¢ĄņĀ£ĒÜ©Ļ│╝ļź╝ Ļ▓ĆņĀĢĒĢśņśĆļŗż(Fig. 4). ļ®ĖĻĘĀņłśņŚÉņä£ ļ░░ņ¢æĒĢ£ ļīĆņĪ░ĻĄ¼ņØś C. acutatum KACC42403 Ēżņ×ÉļŖö ļ░£ņĢäļź╝ ņŗ£ņ×æĒĢśņŚ¼ 24ņŗ£Ļ░äņŚÉ 80.5%, 48ņŗ£Ļ░äņŚÉ 95%Ļ░Ć ļ░£ņĢäĒĢśņśĆļŗż. ļ░śļ®┤ YGB36 ļ░░ņ¢æņĢĪņŚÉņä£ļŖö 48ņŗ£Ļ░äĻ╣īņ¦Ć ņĀäĒśĆ ļ░£ņĢäĒĢśņ¦Ć ļ¬╗ĒĢśņśĆļŗż. YGB36ņØś ņāØĻĘĀņØ┤ ņĀ£Ļ▒░ļÉ£ ļ░░ņ¢æņŚ¼ņĢĪņŚÉ ņä£ļÅä Ēżņ×Éļ░£ņĢä ņ¢ĄņĀ£ĒÜ©Ļ│╝ļŖö ļ¦żņÜ░ ļåÆĻ▓ī ļéśĒāĆļé¼ļŖöļŹ░, 24ņŗ£Ļ░äĻ╣īņ¦Ć ņĀäĒśĆ ļ░£ņĢäĒĢśņ¦Ć ļ¬╗ĒĢśņśĆĻ│Ā 32ņŗ£Ļ░äņŚÉ 12.4%, 48ņŗ£Ļ░äņŚÉļÅä 25.2%Ļ░Ć ļ░£ņĢäĒĢśļŖöļŹ░ ĻĘĖņ│żļŗż. Ēśäļ»ĖĻ▓Įņ£╝ļĪ£ Ļ┤Ćņ░░ĒĢ┤ ļ│┤ļ®┤, ļīĆņĪ░ĻĄ¼ņŚÉņä£ļŖö ņĄ£ņ┤ł Ļ┤Ćņ░░ņŗ£ļČĆĒä░ ļ░£ņĢäĻ┤ĆņØ┤ ļéśĒāĆļé¼Ļ│Ā, 32ņŗ£Ļ░äņŚÉ ļČĆņ░®ĻĖ░Ļ░Ć Ļ┤Ćņ░░ļÉśņŚłņ£╝ļ®░, 48ņŗ£Ļ░äņŚÉļŖö ļŗżņłśņØś ļČĆņ░®ĻĖ░Ļ░Ć Ļ┤Ćņ░░ļÉśņŚłļŗż. ļ░śļ®┤, ļ░░ņ¢æņŚ¼ņĢĪ ņ▓śļ”¼ĻĄ¼ņŚÉņä£ļŖö 32ņŗ£Ļ░äļČĆĒä░ ļ░£ņĢäĻ░Ć ņĪ░ĻĖłņö® ņŗ£ņ×æĒĢśņśĆņ£╝ļéś, Ļ┤Ćņ░░ņØä ņóģļŻīĒĢ£ 48ņŗ£Ļ░äĻ╣īņ¦Ć ņ¢┤ļ¢ż ļČĆņ░®ĻĖ░ļÅä Ļ┤Ćņ░░ļÉśņ¦Ć ņĢŖņĢśļŗż. ļ░░ņ¢æņĢĪ ņ▓śļ”¼ĻĄ¼ņŚÉņä£ļŖö Ļ┤Ćņ░░ņØä ņóģļŻīĒĢ£ 48ņŗ£Ļ░äĻ╣īņ¦Ć ņ¢┤ļ¢ĀĒĢ£ ļ│ĆĒÖöļÅä Ļ┤Ćņ░░ļÉśņ¦Ć ņĢŖņĢśļŗż. B. subtilisĻ░Ć C. acutatumņØś Ēżņ×Éļ░£ņĢäļź╝ ņ¢ĄņĀ£ĒĢ£ļŗżļŖö ņé¼ņŗżņØ┤ ņØ┤ļ»Ė ņŚ¼ļ¤¼ ļģ╝ļ¼ĖņŚÉņä£ ļ│┤Ļ│ĀļÉśņŚłņ£╝ļ®░, YGB36 ņŚÉņä£ļÅä C. acutatumņŚÉ ļīĆĒĢśņŚ¼ Ļ░ĢĒĢ£ ĒÖ£ņä▒ņØ┤ ļéśĒāĆļé¼ļŗż(Lamsal ļō▒, 2012; ┼Įivkovic ļō▒, 2010). ļśÉĒĢ£ B. subtilis CMB32ņØś ļ░░ņ¢æņŚ¼ņĢĪ ņŚÉļŖö iturin A, fengycin, surfactin A ļō▒ņØś lipopetideļōżņØ┤ ĒżĒĢ© ļÉśņ¢┤ ņ׳Ļ│Ā, ņØ┤ļōżņŚÉ ņØśĒĢ£ ĒāäņĀĆļ│æĻĘĀņŚÉņØś ĒĢŁĻĘĀĒÖ£ņä▒ņØ┤ ļ│┤Ļ│ĀļÉśņŚłļŗż(Kim ļō▒, 2010; OngenaņÖĆ Jacques, 2008).
Ļ│ĀņČöĻ│╝ņŗżņŚÉņä£ņØś ĒāäņĀĆļ│æ ļ░£ļ│æņ¢ĄņĀ£.
Ļ│ĀņČö Ļ│╝ņŗżņŚÉ ņāüņ▓śļź╝ ļé┤ņ¢┤ ĒāäņĀĆļ│æĻĘĀņØä ņĀæņóģĒĢ£ Ēøä YGB36 ļ░░ņ¢æņĢĪņØä ĒؼņäØĒĢśņŚ¼ ļČäļ¼┤ĒĢ©ņ£╝ļĪ£ņŹ©, ļ│æ ļ░£ņāØ ņ¢ĄņĀ£ ņĀĢļÅäļź╝ ņĪ░ņé¼ĒĢśņśĆļŗż. ļ░£ļ│æņØ┤ ņ¦äĒ¢ēļÉ£ ļČĆņ£äņŚÉ ņä£ļŖö ņĀäĒśĢņĀüņØĖ ĒāäņĀĆļ│æĻĘĀņØś ļ│æņ¦ĢņØä ĒÖĢņØĖĒĢĀ ņłś ņ׳ņŚłņ£╝ļ®░ ļ░£ņāØ ļ│æļ░śņØś Ļ░£ņłśņÖĆ Ēü¼ĻĖ░ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ļČäņäØĒĢ£ Ļ▓░Ļ│╝, ļīĆņĪ░ĻĄ¼ņØś ļ░£ļ│æļÅäļŖö 97.2%ņ×äņŚÉ ļ░śĒĢ┤ YGB36 ņ▓śļ”¼ĻĄ¼ņŚÉņä£ļŖö ļ░£ļ│æļÅäĻ░Ć 33.3%ļĪ£ ļéśĒāĆļé¼ļŗż(Fig. 5). ļ░®ņĀ£Ļ░ĆļŖö 65.7%ļĪ£ ĒÖśņé░ļÉśņŚłļŗż. ņØ┤ļŖö YGB ļ░░ņ¢æņĢĪ ņØ┤ ĒāäņĀĆļ│æĻĘĀņØś Ēżņ×Éļ░£ņĢä ļ░Å ĻĘĀņé¼ņāØņןņØä ņ¢ĄņĀ£Ē¢łļŹś ņāüĻĖ░ in vitro Ēżņ×Éļ░£ņĢäņ¢ĄņĀ£Ļ▓ĆņĀĢ Ļ▓░Ļ│╝ņÖĆ ņØ╝ņ╣śĒĢśņśĆļŗż.
Ļ│ĀņČö ņóģņ×Éļ░£ņĢä ļ░Å ņāØņ£Īņ┤ēņ¦äĒÜ©Ļ│╝.
YGB36ņØś PGPRĒÜ©Ļ│╝ Ļ▓ĆņĀĢņØä ņ£äĒĢśņŚ¼, ļ░£ņĢäņ£©ņØä Ļ┤Ćņ░░ĒĢśņśĆļŗż. ļ░£ņĢäņ£©ņØĆ Ļ┤Ćņ░░ņØä ņŗ£ņ×æĒĢ£ 9ņØ╝ ņ░©ļČĆĒä░ ņ░©ņØ┤Ļ░Ć ļ░£ņāØĒĢśĻĖ░ ņŗ£ņ×æĒĢśņŚ¼ 10ņØ╝ņ░©ņŚÉ Ļ░Ćņן ļÜ£ļĀĘĒĢ£ ņ░©ņØ┤ļź╝ ļ│┤ņśĆļŗż(Fig. 6A). Ļ┤Ćņ░░ņØä ņóģļŻīĒĢ£ 14ņØ╝ņ░©ņŚÉ ļīĆņĪ░ĻĄ¼ņŚÉņä£ 97.2%, YGB36 ņ▓śļ”¼ĻĄ¼ņŚÉņä£ļŖö 100%ņØś ļ░£ņĢäņ£©ņØä ļ│┤ņśĆļŗż. ļīĆņĪ░ĻĄ¼ ļ░£ņĢäņ£©ņØĆ 9ņØ╝ņ░©ļČĆĒä░ 11ņØ╝ņ░©Ļ╣īņ¦Ć 1.9%, 13.0%, 47.2%ņØĖ ļ░śļ®┤, ņ▓śļ”¼ĻĄ¼ņØś ļ░£ņĢäņ£©ņØĆ 23.1%, 49.1%, 96.3%ļĪ£ ļéśĒāĆļé¼ļŗż. YGB36ļź╝ Ļ│ĀņČö ņóģņ×ÉņŚÉ ņ▓śļ”¼ĒĢśņśĆņØä ļĢī, ļīĆņĪ░ĻĄ¼ņŚÉ ļ╣äĒĢśņŚ¼ ļ│┤ļŗż ļ╣©ļ”¼ ļ░£ņĢäĒĢśĻĖ░ ņŗ£ņ×æņśĆĻ│Ā, 2ņŻ╝ ĒøäņŚÉļŖö YGB36ņ▓śļ”¼ĻĄ¼ņÖĆ ļīĆņĪ░ĻĄ¼ ļ¬©ļæÉņŚÉņä£ ļīĆļČĆļČä ļ░£ņĢäĒĢśņśĆļŗż. Ļ▓░ļĪĀņĀüņ£╝ļĪ£, ļīĆņĪ░ĻĄ¼ņÖĆ YGB36 ņ▓śļ”¼ĻĄ¼ņŚÉņä£ņØś ņĄ£ņóģ ļ░£ņĢäņ£©ņØĆ ļŗżņåī ļ╣äņŖĘĒĢśļéś, YGB36 ņ▓śļ”¼ĻĄ¼ņŚÉņä£ņØś ļ░£ņĢä ņåŹļÅäļŖö ĒśäņĀĆĒ׳ ļ░£ļØ╝ņ¦ä Ļ▓āņØä ņĢī ņłś ņ׳ņŚłļŗż. ļīĆļČĆļČäņØś PGPRņØ┤ ņŚ¼ļ¤¼Ļ░Ćņ¦Ć ĻĖ░ņ×æņ£╝ļĪ£ ļ░£ņĢä ņ┤ēņ¦äĻ│╝ Ļ░ÖņØĆ ņŗØļ¼╝ ņāØņ£ĪņŚÉ ņ£ĀņÜ®ĒĢ£ ĒÜ©Ļ│╝ļź╝ ļéśĒāĆļé┤ļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŖöļŹ░, ņØ╝ļĪĆļĪ£ B. subtilisļŖö indole-3-acetic acid (IAA)ņÖĆ Ļ░ÖņØĆ ņŗØļ¼╝ĒśĖļź┤ļ¬¼ņØä ņāØņé░ĒĢśņŚ¼ ņŗØļ¼╝Ļ│╝ņØś ņāüĒśĖņ×æņÜ®ņØä ĒĢ£ļŗż(Swain ļō▒, 2007). ņäĖĻĘĀņØś IAAļŖö ņŗØļ¼╝ņäĖĒżņØś ļČäņŚ┤Ļ│╝ ĒÖĢņן(expansion)ņØä ņ┤ēņ¦äĒĢśņŚ¼ ļ┐īļ”¼Ļ░Ć ņČ£ĒśäĒĢĀ ņłś ņ׳Ļ▓ī ĒĢ©ņ£╝ļĪ£ņŹ© ņóģņ×É Ē£┤ļ®┤ņØä ĒāĆĒīīĒĢśļŖöļŹ░ ļÅäņøĆņØä ņżä ņłś ņ׳ļŗżļŖö ņé¼ņŗżņØ┤ ņל ņĢīļĀżņĀĖ ņ׳ļŗż(Spaepen ļō▒, 2007). YGB36ņØä ņ▓śļ”¼ĒĢśņŚ¼ ļéśĒāĆļé£ Ļ│ĀņČöņóģņ×ÉņØś ļ░£ņĢäņ┤ēņ¦äļÅä ņØ┤ņÖĆ Ļ░ÖņØĆ ĻĖ░ņ×æņØ┤ Ļ┤ĆļĀ©ņØ┤ ņ׳ņØä Ļ▓āņ£╝ļĪ£ ĒīÉļŗ©ļÉ£ļŗż.
ņóģņ×É ļ░£ņĢä ņØ┤ĒøäņØś ņ┤łĻĖ░ ņāØņ£ĪĒÜ©Ļ│╝ Ļ▓ĆņĀĢņØä ņ£äĒĢ┤ Ēīīņóģ Ēøä 15ņØ╝ ņ░©ņŚÉ ļ┐īļ”¼ ĻĖĖņØ┤ļź╝ ņĖĪņĀĢĒĢśņśĆļŗż. ļīĆņĪ░ĻĄ¼ļŖö ĒÅēĻĘĀ 39.1 mmņØĖ ļ░śļ®┤ ņ▓śļ”¼ĻĄ¼ņŚÉņä£ļŖö ĒÅēĻĘĀ 53.6 mmļĪ£, LSD test Ļ▓░Ļ│╝, P=0.019ļĪ£ ļīĆņĪ░ ĻĄ¼ņÖĆ ņ▓śļ”¼ĻĄ¼ ņé¼ņØ┤ņŚÉļŖö ņ£ĀņØśņä▒ņØ┤ ņ׳ļŖö Ļ▓āņ£╝ļĪ£ ļéśĒāĆļé¼ļŗż(Fig. 6B). ļö░ļØ╝ņä£, YGB36ņØĆ Ļ│ĀņČö ņóģņ×É ļ░£ņĢä ņØ┤ĒøäņØś ņ┤łĻĖ░ ņāØņ£ĪņØä ņ┤ēņ¦äĒĢśļŖö Ļ▓āņ£╝ļĪ£ ĒīÉļŗ©ļÉśņŚłļŗż.
Ēīīņóģ Ēøä 58ņØ╝ņØ┤ ņ¦Ćļéś ņ¦ĆņāüļČĆ ĻĖĖņØ┤ļź╝ ņĖĪņĀĢĒĢśņśĆļŗż. ļīĆņĪ░ĻĄ¼ņÖĆ ņ▓śļ”¼ĻĄ¼Ļ░äņØś ņ¦ĆņāüļČĆ ņāØņ£ĪņØĆ ņ£ĪņĢłņ£╝ļĪ£ļÅä ĻĄ¼ļ│äļÉĀ ļ¦īĒü╝ ņ░©ņØ┤Ļ░Ć ņ╗Ėņ£╝ļ®░ ņĖĪņĀĢĻ▓░Ļ│╝, ļīĆņĪ░ĻĄ¼ņØś ņ¦ĆņāüļČĆ ĻĖĖņØ┤ļŖö ĒÅēĻĘĀ 25.0 cmņØĖ ļ░śļ®┤, ņ▓śļ”¼ĻĄ¼ņØś ĒÅēĻĘĀ ĻĖĖņØ┤ļŖö 28.8 cmņśĆļŗż. LSD test Ļ▓░Ļ│╝, P<0.001ļĪ£ ļīĆņĪ░ĻĄ¼ņÖĆ ņ▓śļ”¼ĻĄ¼ ņé¼ņØ┤ņŚÉļŖö ņ£ĀņØśņä▒ņØ┤ ļ¦żņÜ░ ļåÆņØĆ Ļ▓āņ£╝ļĪ£ ļéśĒāĆļé¼ļŗż (Fig. 6C). ļö░ļØ╝ņä£, YGB36ņØś Ļ│ĀņČö ņāØņ£Īņ┤ēņ¦ä ĒÜ©Ļ│╝Ļ░Ć ļ¦żņÜ░ ļåÆņØĆ Ļ▓āņ£╝ļĪ£ ĒīÉļŗ©ļÉśņŚłļŗż.
ĒÖöĒĢÖļåŹņĢĮņŚÉ ļīĆĒĢ£ ņĀĆĒĢŁņä▒ Ļ▓ĆņĀĢ.
ņāØļ¼╝ĒĢÖņĀü ļ░®ņĀ£ņĀ£ļź╝ ĒÖöĒĢÖļåŹ ņĢĮĻ│╝ Ēś╝ņÜ® Ēś╣ņØĆ ĻĄÉĒśĖ ņé┤ĒżĒĢśņśĆņØä ļĢī ņä£ļĪ£ ņāüņČ®ņ×æņÜ®ņØä ĒĢśļŖöņ¦Ć ņĢīņĢäļ│┤ĻĖ░ ņ£äĒĢ┤ ļŗżņØīĻ│╝ Ļ░ÖņØĆ ņŗżĒŚśņØä ņłśĒ¢ēĒĢśņśĆļŗż. ņāüņÜ®ĒÖöļÉśņ¢┤ ņ׳ļŖö ņé┤ĻĘĀņĀ£ 21ņóģ ļ░Å ņé┤ņČ®ņĀ£ 14ņóģņØä ņĀ£ņĪ░ņé¼ņØś ņ¦Ćņ╣©ņØä ĻĖ░ņżĆņ£╝ļĪ£ ļ░śļ¤ē, ņĀĢļ¤ē ļ░Å ļ░░ļ¤ēņ£╝ļĪ£ ņĀ£ņĪ░ĒĢśņŚ¼ ņØ┤ņŚÉ ļīĆĒĢ£ YGB36ņØś ņāØņ£ĪņŚÉ ņØś ņśüĒ¢źņØä ĒÖĢņØĖĒĢśņśĆļŗż(Table 1). ņ┤Ø 21ņóģņØś ņé┤ĻĘĀņĀ£ ņżæņŚÉņä£ pyraclostrobin ļ░Å tebuconazole ļō▒ņØś ņøÉņĀ£ļź╝ ĒżĒĢ©ĒĢ£ ņé┤ĻĘĀņĀ£ 11ņóģņØĆ YGB36ņØś ņāØņ£ĪņŚÉ ņĀäĒśĆ ņ¦ĆņןņØä ņŻ╝ņ¦Ć ņĢŖņĢśņ£╝ļ®░, streptomycin Ļ│╝ validamycin-AņØś ĒĢ®ņĀ£ņØś Ļ▓ĮņÜ░ ņĀĢļ¤ē ņØ┤ņāüņŚÉņä£ļ¦ī ņāØņ£Ī ņĀĆĒĢ┤Ļ░Ć ļéśĒāĆļé¼ļŗż. ĒĢ£ĒÄĖ, ņŗżĒŚśņŚÉ ņé¼ņÜ®ĒĢ£ ņé┤ņČ®ņĀ£ 14ņóģņØĆ YGB36ņØś ņāØņ£Ī ņŚÉ ņĀäĒśĆ ņśüĒ¢źņØä ņŻ╝ņ¦Ć ņĢŖņĢśļŗż. ļö░ļØ╝ņä£, YGB36Ļ│╝ ņé┤ĻĘĀņĀ£ņØś Ēś╝ņÜ® ņŚ¼ļČĆļŖö ņé┤ĻĘĀņĀ£ ņóģļźśņŚÉ ļö░ļØ╝ ļŗżļź┤ļ»ĆļĪ£, Ēś╝ņÜ® ņŗ£ ņŻ╝ņØśĒĢśļŖö Ļ▓āņØ┤ ļ░öļ×īņ¦üĒĢśļ®░, ņé┤ņČ®ņĀ£ņÖĆņØś Ēś╝ņÜ®ņŚÉņä£ļŖö ņØ┤ļź╝ Ļ│ĀļĀżĒĢśņ¦Ć ņĢŖņĢäļÅä ļÉĀ Ļ▓āņ£╝ļĪ£ ĒīÉļŗ©ļÉ£ļŗż. ļŗżļ¦ī, ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ņé¼ņÜ®ĒĢ£ ņé┤ĻĘĀņĀ£ ļ░Å ņé┤ņČ®ņĀ£Ļ░Ć Ēśäņ×¼ ņāüņÜ®ĒÖöļÉ£ ļ¬©ļōĀ ĒÖöĒĢÖļåŹņĢĮņØä ļŗżļŻ©ņ¦ĆļŖö ļ¬╗Ē¢łņ£╝ļ»ĆļĪ£ ņāØļ¼╝ĒĢÖņĀü ņĀ£ņĀ£ Ļ░£ļ░£ ņŗ£ ļŗżņ¢æĒĢ£ ĒÖöĒĢÖļåŹņĢĮ ļ░Å ļŗżļźĖ ļ»ĖņāØļ¼╝ņĀ£ņÖĆņØś Ēś╝ņÜ® ņŚ¼ļČĆņŚÉ ļīĆĒĢ£ ņŚ░ĻĄ¼Ļ░Ć ĒĢäņÜöĒĢĀ Ļ▓āņ£╝ļĪ£ ĒīÉļŗ©ļÉ£ļŗż.
ņ×ÉņŚ░Ļ│äņŚÉļŖö ņĢäņ¦ü ĒāÉņāēļÉśņ¦Ć ņĢŖņØĆ ņ£ĀņÜ®ņ×ÉņøÉņØ┤ ļ¦ÄņØ┤ ņĪ┤ņ×¼ĒĢśĻ│Ā ņ׳ņ£╝ļ®░, ņ╣£ĒÖśĻ▓Į ņāØļ¼╝ļåŹņĢĮņ£╝ļĪ£ņØś Ļ░£ļ░£Ļ░ĆļŖźņä▒ņØ┤ ļ¦żņÜ░ ļåÆņØĆ ļ»ĖņāØļ¼╝ņØä ĒāÉņāēĒĢśņŚ¼ ņ£ĀņÜ®ņ×ÉņøÉņ£╝ļĪ£ņä£ ĒÖ£ņÜ®ĒĢśļŖö ņŗ£ļÅäļŖö ņśżļלņĀäļČĆĒä░ ņØ┤ļŻ©ņ¢┤ņĀĖ ņśżĻ│Ā ņ׳ļŗż. ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ĒāÉņāēļÉ£ YGB36ņØĆ Ļ░ĢĒĢ£ ĒĢŁĻĘĀĒÖ£ ņä▒Ļ│╝ ļŹöļČłņ¢┤ ņØĖņé╝ļ┐īļ”¼ņŹ®ņØīļ│æĻĘĀ(C. destructans)Ļ│╝ ĒāäņĀĆļ│æĻĘĀ(C. acutatum, C. gloeosporioides)Ļ│╝ Ļ░ÖņØ┤ ļäōņØĆ ĒĢŁĻĘĀņŖżĒÄÖĒŖĖļ¤╝ņØä ļ│┤ņ£ĀĒĢśĻ│Ā ņ׳ļŗż. Ļ▓īļŗżĻ░Ć siderophore ņāØņé░ ļ░Å ļé£ņÜ®ņä▒ ņØĖņé░ņŚ╝ ļČäĒĢ┤ļŖź ļśÉĒĢ£ ļ│┤ņ£ĀĒĢśĻ│Ā ņ׳ņ¢┤ ņØ┤ļ¤¼ĒĢ£ ĒŖ╣ņ¦ĢļōżņØ┤ ņŗØļ¼╝ņāØņןņŚÉ ņ¦üņĀæņĀüņØĖ ļÅäņøĆņØä ņżä ņłś ņ׳ņØä Ļ▓āņ£╝ļĪ£ ĻĖ░ļīĆļÉ£ļŗż. ņŗżņĀ£ļĪ£ YGB36ņØ┤ Ļ│ĀņČöņŚÉņä£ Ļ░Ćņן ļ¼ĖņĀ£Ļ░Ć ļÉśĻ│Ā ņ׳ļŖö ĒāäņĀĆļ│æĻĘĀņØä ņ¢ĄņĀ£ĒĢśĻ│Ā, Ļ│ĀņČö ņóģņ×É ļ░£ņĢä ļ░Å ņāØņ£Īņ┤ēņ¦äĻ╣īņ¦Ć ĻĖŹņĀĢņĀüņØĖ ĒÜ©Ļ│╝ļź╝ ļ│┤ņ£ĀĒĢśĻ│Ā ņ׳ņØīņØ┤ ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ĒÖĢņØĖļÉśņŚłļŗż. ņĢäņÜĖļ¤¼ ņŗżņĀ£ ļåŹĻ░ĆņŚÉņä£ ņŻ╝ļĪ£ ņé¼ņÜ®ĒĢśĻ│Ā ņ׳ļŖö ļŗżņ¢æĒĢ£ ĒÖöĒĢÖņĢĮņĀ£ņÖĆņØś Ēś╝ņÜ® Ļ░ĆļŖźņä▒ļÅä ņĀ£ņŗ£ĒĢśņśĆļŗż. ĒĢ£ĒÄĖ YGB36ņØĆ B. subtilisņóģņ£╝ļĪ£ņä£, ĒĢ┤ļŗ╣ ņóģņØĆ ņĢłņĀäņä▒ņØ┤ ņ×ģņ”ØļÉśņ¢┤ ņØ┤ļ»Ė ņśżļ×£ ĻĖ░Ļ░ä ņé¼ņÜ®ļÉśņ¢┤ ņś© ĻĘĀņŻ╝ņØ┤ļ»ĆļĪ£ ņāØļ¼╝ļåŹņĢĮņ£╝ļĪ£ņä£ņØś Ļ░£ļ░£ņŚÉ ĒĢ£ņĖĄ ņÜ®ņØ┤ĒĢśļŗż. ņØ┤ļĪ£ņŹ© YGB36ņØĆ Ļ│ĀņČöĒāäņĀĆļ│æ ņ¢ĄņĀ£ ļ░Å ņŗØļ¼╝ņāØņ£Īņ┤ēņ¦äņØä ņ£äĒĢ£ ņ╣£ĒÖśĻ▓ĮņĀ£ņĀ£ļĪ£ņä£ņØś ļåÆņØĆ ĒÖ£ņÜ®Ļ░ĆļŖźņä▒ņØä ļ│┤ņ£ĀĒĢśĻ│Ā ņ׳ļŖö Ļ▓āņ£╝ļĪ£ ĒīÉļŗ©ļÉ£ļŗż.

 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






