


ļöĖĻĖ░(Fragaria spp.)ļŖö ņןļ»ĖĻ│╝ņŚÉ ņåŹĒĢśļŖö ļŗżļģäņāØ ņŗØļ¼╝ļĪ£ ņĀä ņäĖĻ│äņĀüņ£╝ļĪ£ 20ņŚ¼ ņóģ(species)Ļ│╝ ņłśļ¦ÄņØĆ ĻĄÉļ░░ņóģ(hybrids) ļ░Å ĒÆłņóģ(cultivars)ņØ┤ ņĪ┤ņ×¼ĒĢ£ļŗż. ņ×¼ļ░░ņóģ ļöĖĻĖ░ņØĖ Fragaria ├Ś ananassa ļŖö F. virginiana (ļČüņĢäļ®öļ”¼ņ╣┤ ņ£Āļל)ņÖĆ F. chiloensis (ļé©ņĢäļ®öļ”¼ņ╣┤ ņ£Āļל)ņØś ĻĄÉļ░░ņóģņ£╝ļĪ£ 1750ļģäļīĆ ņ£Āļ¤ĮņŚÉņä£ ņ£ĀļלļÉśņ¢┤ ņ¦ĆĻĖłĻ╣īņ¦Ć ņĀä ņäĖĻ│äņĀüņ£╝ļĪ£ Ļ░Ćņן ļ¦ÄņØ┤ ņ×¼ļ░░ļÉśĻ│Ā ņ׳ņ£╝ļ®░ ļöĖĻĖ░ ņ£ĪņóģņØś ĻĖ░ļ│ĖĒśĢņ£╝ļĪ£ ņØ┤ņÜ®ļÉśĻ│Ā ņ׳ļŗż(MartinĻ│╝ Tzanetakis, 2006; Staudt, 1962). ĻĄŁļé┤ņŚÉ ļöĖĻĖ░(F. ├Ś ananassa)Ļ░Ć ļÅäņ×ģļÉ£ Ļ▓āņØĆ 1900ļģäļīĆ ņ┤łĻĖ░ļĪ£ 2000ļģäļīĆ ņ┤łļ░śĻ╣īņ¦Ć ņŻ╝ļĪ£ ņØ╝ļ│Ė ĒÆłņóģņØ┤ ņ×¼ļ░░ļÉśņŚłņ£╝ļéś 1990ļģä Ēøäļ░ś ņØ┤Ēøä ĻĄŁļé┤ ņ£Īņä▒ ĒÆłņóģ Ļ░£ļ░£ņØ┤ ĒÖ£ļ░£Ē׳ ņØ┤ļŻ©ņ¢┤ņĀĖ Ēśäņ×¼ ĻĄŁņé░ ļöĖĻĖ░ ĒÆłņóģņØś ļ│┤ĻĖēļźĀņØĆ 93% ņØ┤ņāüņØä ņ░©ņ¦ĆĒĢśĻ│Ā ņ׳ļŗż.
ļöĖĻĖ░ļŖö ņśüņ¢æļ▓łņŗØņŚÉ ņØśĒĢ┤ ļŗżļģäņāØņ£╝ļĪ£ ņ×¼ļ░░ļÉśļ»ĆļĪ£ ļ░öņØ┤ļ¤¼ņŖżņŚÉ Ļ░ÉņŚ╝ļÉĀ Ļ▓ĮņÜ░ ļ░öņØ┤ļ¤¼ņŖżĻ░Ć ņČĢņĀüļÉśņ¢┤ ņāØņ£Ī ļ░Å ņłśļ¤ēņŚÉ ņśüĒ¢źņØä ļ░øņØä ņłś ņ׳ļŗż. ļöĖĻĖ░ņŚÉ ļ░£ņāØĒĢ£ļŗżĻ│Ā ļ│┤Ļ│ĀļÉ£ ļ░öņØ┤ļ¤¼ņŖżļŖö ņĢĮ 30ņŚ¼ ņóģņØ┤ļ®░ ĻĘĖ ņżæ Strawberry mild yellow edge virus (SMYEV, genus Potexvirus), Strawberry mottle virus (SMoV, family Seco viridae, genus not assigned), Strawberry crinkle virus (SCV, genus Cytorhabdovirus), Strawberry vein banding virus (SVBV, genus Caulimovirus)Ļ░Ć Ļ▓ĮņĀ£ņĀüņ£╝ļĪ£ Ļ░Ćņן ņżæņÜöĒĢ£ ļ░öņØ┤ļ¤¼ņŖżļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż(Franova, 2001; MartinĻ│╝ Tzanetakis, 2006; MellorņÖĆ Krczal, 1987; SpiegelĻ│╝ Martin, 1998). ņØ┤ļōż ļ░öņØ┤ļ¤¼ņŖżļŖö ļ¬©ļæÉ ņ¦äļö¦ļ¼╝ņŚÉ ņØśĒĢ┤ ņĀäļ░śļÉśļŖö ļ░öņØ┤ļ¤¼ņŖżļĪ£ ļöĖĻĖ░ ĒÆłņóģ ļ░Å ļ░öņØ┤ļ¤¼ņŖż Ļ│äĒåĄ(strain)ņŚÉ ļö░ļØ╝ ļ│æņøÉņä▒ņŚÉ ņ░©ņØ┤Ļ░Ć ņ׳ņ¦Ćļ¦ī F. vesca (ņĢ╝ņāØļöĖĻĖ░)ļéś F. virginianaņÖĆ Ļ░ÖņØĆ Ļ░Éņłśņä▒ ĒÆłņóģņØś Ļ▓ĮņÜ░ Ļ│╝ņŗżņØś Ē¢źņØä ļ¢©ņ¢┤ļ£©ļ”¼Ļ│Ā ņāē ļ░£ĒśäņØ┤ ņל ļÉśņ¦Ć ņĢŖņ£╝ļ®░ ļ¤░ļäł(runner) ņāØņé░ņØä ņĀĆĒĢśņŗ£Ēé©ļŗżĻ│Ā ļ│┤Ļ│ĀļÉśĻ│Ā ņ׳ļŗż(Frazier, 1974; FreemanĻ│╝ Mellor, 1962; HornĻ│╝ Carver, 1962). ĒĢ£ĒÄĖ, ņ×¼ļ░░ļöĖĻĖ░(F. ├Ś ananassa)ņØś ņ£Īņä▒ĒÆłņóģ ļīĆļČĆļČäņØĆ ļ░öņØ┤ļ¤¼ņŖżņŚÉ ļé┤ņä▒(tolerant)ņØ┤ ņ׳ļŗżĻ│Ā ņĢīļĀżņĀĖ ņ׳ņ¢┤ ļŗ©ņØ╝Ļ░ÉņŚ╝ņŚÉņä£ļŖö ļ│æņ¦ĢņØä ļÅÖļ░śĒĢśņ¦Ć ņĢŖĻ▒░ļéś Ēö╝ĒĢ┤Ļ░Ć ņĢĮĒĢśņ¦Ćļ¦ī 2ņóģ ņØ┤ņāüņØś ļ░öņØ┤ļ¤¼ņŖżĻ░Ć ļ│ĄĒĢ®Ļ░ÉņŚ╝ņØ┤ ļÉĀ Ļ▓ĮņÜ░ ļöĖĻĖ░ ņłśļ¤ēņØä ĻĖēĻ▓®Ē׳ ņĀĆĒĢśņŗ£ĒéżĻ│Ā Ēć┤ĒÖöļź╝ ņ£ĀļÅäĒĢ£ļŗż(SpiegelĻ│╝ Martin, 1998). ņØ┤ļ¤¼ĒĢ£ ņØ┤ņ£ĀļĪ£ ļöĖĻĖ░ ņ×¼ļ░░ ņŗ£ ļ░öņØ┤ļ¤¼ņŖżļź╝ ļ¦żĻ░£ĒĢśļŖö ņ¦äļö¦ļ¼╝ ļ░®ņĀ£ļŖö ļ¦żņÜ░ ņżæņÜöĒĢśļŗż. ļöĖĻĖ░ ļ░öņØ┤ļ¤¼ņŖżļź╝ ņĀäļ░śĒĢśļŖö ļīĆĒæ£ņĀüņØĖ ņ¦äļö¦ļ¼╝ņØĆ Chaetosiphon fragaefoliiļĪ£ SMYEV, SCV, SVBVļŖö Chaetosiphon spp.ņŚÉ ņØśĒĢ┤ņä£ļ¦ī ņĀäļ░śļÉśļŖö ļ░śļ®┤, SMoVņØś Ļ▓ĮņÜ░ Chaetosiphon spp.ņÖĆ Aphis gossypii (ļ¬®ĒÖöņ¦äļö¦ļ¼╝)ņŚÉ ņØśĒĢ┤ ņĀäļ░śļÉ£ļŗż(Krczal, 1982; MartinĻ│╝ Tzanetakis, 2006). ĻĄŁļé┤ņØś Ļ▓ĮņÜ░ 2007-2008ļģäņŚÉ ļöĖĻĖ░ ļ░öņØ┤ļ¤¼ņŖżļ│æ ņĪ░ņé¼Ļ░Ć ņØ┤ļŻ©ņ¢┤ņ¦ä ļ░ö ņ׳ņ£╝ļéś(Choi ļō▒, 2009), ĻĄŁļé┤ ņ£Īņä▒ ĒÆłņóģņØ┤ ļäÉļ”¼ ļ│┤ĻĖēļÉ£ ņØ┤Ēøä ļöĖĻĖ░ ĒżņןņŚÉņä£ ļ░öņØ┤ļ¤¼ņŖżļ│æ ņĪ░ņé¼Ļ░Ć ņØ┤ļŻ©ņ¢┤ņ¦ä ļ░ö ņŚåņ¢┤ ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ļöĖĻĖ░ņŚÉ Ļ░Ćņן ļ¼ĖņĀ£Ļ░Ć ļÉśļŖö ņ¦äļö¦ļ¼╝ ņĀäļ░ś ļöĖĻĖ░ ļ░öņØ┤ļ¤¼ņŖż 4ņóģ(SMYEV, SMoV, SCV, SVBV)ņØä ļīĆņāüņ£╝ļĪ£ ļöĖĻĖ░ ņŻ╝ņÜö ņ×¼ļ░░ņ¦ĆņŚŁ(ņ¦äņŻ╝, ļģ╝ņé░, Ļ│Āņä▒, ĒĢ©ņ¢æ, ņł£ņ░Į, ĒÅēņ░Į, Ļ░Ģļ”ē)Ļ│╝ ĻĄŁļé┤ ņ£Īņä▒ ņŻ╝ņÜö ņ×¼ļ░░ ĒÆłņóģ(ļ¦żĒ¢ź, ņäżĒ¢ź, ĻĖłņŗż, ņŗ╝ĒāĆ, ļ¼┤ĒĢś, ņןĒĢś, Ļ│ĀĒĢś, ņŻĮĒ¢ź)ņØä ļīĆņāüņ£╝ļĪ£ ļ░öņØ┤ļ¤¼ņŖżļ│æ ļ░£ņāØ ņŚ¼ļČĆļź╝ ņĪ░ņé¼ĒĢśņśĆļŗż.
ĻĄŁļé┤ ņ£Īņä▒ ļöĖĻĖ░ ĒÆłņóģņØś ņ¦äļö¦ļ¼╝ ņĀäļ░ś ļöĖĻĖ░ ļ░öņØ┤ļ¤¼ņŖż ļ░£ņāØņĪ░ņé¼
4ņóģņØś ņ¦äļö¦ļ¼╝ ņĀäļ░ś ļöĖĻĖ░ ļ░öņØ┤ļ¤¼ņŖżļź╝ ņ¦äļŗ©ĒĢśĻĖ░ ņ£äĒĢ┤ ļ©╝ņĀĆ Ļ░ü ļ░öņØ┤ļ¤¼ņŖżņØś ĒŖ╣ņØ┤ ĒöäļØ╝ņØ┤ļ©Ėļź╝ ņĀ£ņ×æĒĢśņśĆļŗż. ņĀĢĒÖĢĒĢ£ ņ¦äļŗ©ņØä ņ£äĒĢ┤ GenBank databaseļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ Ļ░ü ļ░öņØ┤ļ¤¼ņŖżņØś ļŗżņ¢æĒĢ£ ļČäļ”¼ņŻ╝(isolate)ņØś ņŚ╝ĻĖ░ņä£ņŚ┤ņØä ļ╣äĻĄÉĒĢśņŚ¼ Ļ░Ćņן ļ│┤ņĪ┤ņØ┤ ņל ļÉśņ¢┤ņ׳ļŖö Ļ░ü ļ░öņØ┤ļ¤¼ņŖżļ│ä ņ£ĀņĀäņ×É ņŚ╝ĻĖ░ņä£ņŚ┤ņØä ļ░öĒāĢņ£╝ļĪ£ ĒöäļØ╝ņØ┤ļ©Ėļź╝ ņĀ£ņ×æĒĢśņśĆļŗż(Table 1). 2018ļģä-2019ļģäĻ╣īņ¦Ć ļöĖĻĖ░ ņ×¼ļ░░ ļåŹĻ░Ć ļ░Å ņøÉļ¼śņןņŚÉņä£ ņØ┤ņāü ņ”ØņāüņØä ļ│┤ņØ┤ļŖö ņ×Ä ņŗ£ļŻīņÖĆ ļ¼┤ļ│æņ¦Ģ ņŗ£ļŻīļź╝ ļ¼┤ņ×æņ£äļĪ£ ņ▒äņ¦æĒĢśņŚ¼ ņ┤Ø 918ņĀÉ(ļ¦żĒ¢ź 322ņĀÉ, ņäżĒ¢ź 183ņĀÉ, ĻĖłņŗż 169ņĀÉ, ņŗ╝ĒāĆ 42ņĀÉ, ļ¼┤ĒĢś88ņĀÉ, ņןĒĢś 51ņĀÉ, ĻĖ░ĒāĆ[Ļ│ĀĒĢś, ņŻĮĒ¢ź] 63ņĀÉ)ņŚÉ ļīĆĒĢ┤ ĻĖ░ņĪ┤ ļ░®ļ▓ĢĻ│╝ ļÅÖņØ╝ĒĢśĻ▓ī ĒĢĄņé░ņØä ņČöņČ£ĒĢśĻ│Ā ņ£ĀņĀäņ×É ņ¦äļŗ©ņØä ņŗżņŗ£ĒĢśņśĆļŗż(Kwon ļō▒, 2018). ņ¦äļŗ© Ļ▓░Ļ│╝ ņ▒äņ¦æ ņ¦ĆņŚŁņŚÉ ļö░ļźĖ ņ£ĀņØśņä▒ņØĆ ņŚåņŚłņ£╝ļ®░ SMYEVĻ░Ć ļ¦żĒ¢ź, ņäżĒ¢ź, ĻĖłņŗżņŚÉņä£ Ļ░üĻ░ü 0.9%, 0.5%, 0.6%, SMoVĻ░Ć ļ¦żĒ¢ź, ņäżĒ¢ź, ĻĖłņŗż, ņŗ╝ĒāĆņŚÉņä£ Ļ░üĻ░ü 0.3%, 1.0%, 1.1%, 2.4%ņØś ļ░£ņāØļźĀļĪ£ Ļ▓ĆņČ£ļÉśņŚłĻ│Ā SCVņÖĆ SVBVļŖö ņ¢┤ļ¢ż ĒÆłņóģņŚÉņä£ļÅä Ļ▓ĆņČ£ļÉśņ¦Ć ņĢŖņĢśļŗż(Table 2). ĒĢ£ĒÄĖ, ļ░öņØ┤ļ¤¼ņŖżĻ░Ć Ļ▓ĆņČ£ļÉ£ ņŗ£ļŻīņŚÉņä£ ļÜ£ļĀĘĒĢ£ ļ░öņØ┤ļ¤¼ņŖż ņ”ØņāüņØĆ ļ░£Ļ▓¼ļÉśņ¦Ć ņĢŖņĢśĻ│Ā ĒĢ£ ņŗ£ļŻīņŚÉņä£ 2ņóģņØ┤ ĒĢ©Ļ╗ś Ļ▓ĆņČ£ļÉ£ ļ│ĄĒĢ®Ļ░ÉņŚ╝ ņŗ£ļŻīļŖö ņŚåņŚłļŗż(data not shown). ĻĄŁļé┤ ņ£Īņä▒ ļöĖĻĖ░ņŚÉņä£ SMYEVņŚÉ ļīĆĒĢ£ ĒŖ╣ņä▒ ļ░Å ĻĄŁļé┤ ļČäļ”¼ņŻ╝ņØś ņ£ĀņĀäņ×É ļČäņäØņØĆ ņĄ£ĻĘ╝ņŚÉ ļ│┤Ļ│ĀļÉ£ ļ░śļ®┤(Cho ļō▒, 2011; Kwon ļō▒, 2018), SMoVņŚÉ ļīĆĒĢ£ ĻĄŁļé┤ ļČäļ”¼ņŻ╝ņŚÉ ņ£ĀņĀäņĀü ļČäņäØņØĆ ļŗżļżäņ¦ä ļ░ö ņŚåņ¢┤ ņ”ØĒÅŁņé░ļ¼╝ņŚÉ ļīĆĒĢ£ ņŚ╝ĻĖ░ņä£ņŚ┤ ļČäņäØņØä ņČöĻ░ĆļĪ£ ņ¦äĒ¢ēĒĢśņśĆļŗż.
Table┬Ā1
List of primers used to detect four aphid-transmitted strawberry viruses
| Virus | Primer | Sequence (5ŌĆÖŌåÆ3ŌĆÖ) | Reference or GenBank database | Gene | Size (bp) |
|---|---|---|---|---|---|
| SMYEV | CF | ACAATCGCCCTGGTCAGTAATTCC | Kwon et al. (2018) | CP | 729 |
| CR | TGGAGCTGGGTAGAAGAAGTGAGA | ||||
| SMoV | 3UF | CGACAGTTCTCTATGTAGGACACC | AJ311875a, AJ311876b, KU177218a, KU177219b | 3' UTR | 782 |
| 3UR | CATCTATCTAAAGTTAAGTCTACA | KU200454b, KU200457b, KU200459b, KU200461b | |||
| SCV | RdF | ATATCCGGATTTGAGAACA | AY250986, AY331389, AY331386, AY331390, | RdRp | 527 |
| RdR | CCTTAACATTGGTGGCAGAC | AY331385, AY331387, AY331388 | |||
| SVBV | CF | AGAGAAAGACTCGAACAATG | HE681085, KT250632, MH894295, X97304, | CP | 959 |
| CR | CCATATTGTGTTTCCGGTGA | KR080547, KP311681, KX950836, KX787430 | |||
Table┬Ā2
Incidence of four aphid-transmitted strawberry viruses in major Korean cultivars during 2018-2019
| Virus | Fragaria ananassa cv. | ||||||
|---|---|---|---|---|---|---|---|
|
|
|||||||
| Maehyang | Seolhyang | Geumsil | Santa | Mooha | Jangha | Othersa | |
| SMYEV | 3/322 | 1/183 | 1/169 | 0/42 | 0/88 | 0/51 | 0/63 |
| SMoV | 1/322 | 2/183 | 2/169 | 1/42 | 0/88 | 0/51 | 0/63 |
| SCV | 0/322 | 0/183 | 0/169 | 0/42 | 0/88 | 0/51 | 0/63 |
| SVBV | 0/322 | 0/183 | 0/169 | 0/42 | 0/88 | 0/51 | 0/63 |
SMoV ĻĄŁļé┤ ļČäļ”¼ņŻ╝ņØś 3ŌĆ▓ untranslated region (UTR) ņŚ╝ĻĖ░ņä£ņŚ┤ņØś ņ£ĀņĀäĒĢÖņĀü ĒŖ╣ņä▒ ļ░Å Ļ│äĒåĄ ļČäņäØ
ļ¦żĒ¢ź, ņäżĒ¢ź, ĻĖłņŗż, ņŗ╝ĒāĆņŚÉņä£ ļČäļ”¼ļÉ£ ņ┤Ø 6Ļ░£ņØś SMoV 3ŌĆ▓ UTR ņ”ØĒÅŁņé░ļ¼╝ņØĆ Kwon ļō▒(2018)ņØś ļ░®ļ▓ĢĻ│╝ ļÅÖņØ╝ĒĢśĻ▓ī ņŚ╝ĻĖ░ņä£ņŚ┤ņØä ļČäņäØĒĢśĻ│Ā GenBankņŚÉ ļō▒ļĪØĒĢśņśĆļŗż: MaeH (MN520311), SH-P (MN520312), SH-S(MN520313), GS-G (MN520309), GS-S (MN520310), ST(MN520314). 6Ļ░£ņØś ĻĄŁļé┤ ļČäļ”¼ņŻ╝ņÖĆ ĒĢ©Ļ╗ś SMoV Ļ░ÉņŚ╝ņŻ╝ļĪ£ ĒÖĢļ│┤ĒĢśĻ│Ā ņ׳ļŖö ļ»ĖĒÖŹ ĒÆłņóģ(MH)ļÅä ļČäņäØņŚÉ ĒĢ©Ļ╗ś ĒżĒĢ©ņŗ£ņ╝░ļŗż(Table 3). 7Ļ░£ņØś ĻĄŁļé┤ ļČäļ”¼ņŻ╝ņÖĆ ĻĖ░ņĪ┤ņŚÉ ļ│┤Ļ│ĀļÉ£ ļČäļ”¼ņŻ╝ 1134 RNA1Ļ│╝ RNA2ņØś 3ŌĆ▓ UTR 782 nt ņä£ņŚ┤ņØä ClustalX ĒöäļĪ£ĻĘĖļשņØä ņØ┤ņÜ®ĒĢ┤ ļ╣äĻĄÉ ļČäņäØĒ¢łļŗż(Table 3). ĻĘĖ Ļ▓░Ļ│╝, 7Ļ░£ņØś ĻĄŁļé┤ ļČäļ”¼ņŻ╝ ļ¬©ļæÉ Ļ│ĄĒåĄņĀüņ£╝ļĪ£ ļČäļ”¼ņŻ╝ 1134ņŚÉ ļ╣äĒĢ┤ ņŚ╝ĻĖ░Ļ░Ć 3Ļ░£(TCT, at position 672-675) ļŹö ļ¦ÄņĢśņ£╝ļ®░ ST-P ļČäļ”¼ņŻ╝ļŖö ĒĢśļéśņØś ņŚ╝ĻĖ░(C, at position 312)Ļ░Ć ļŹö ņ׳ļŖö Ļ▓āņØä ņĢī ņłś ņ׳ņŚłļŗż. ļČäļ”¼ņŻ╝ 1134ņØś RNA1Ļ│╝ 2ņØś ļ╣äĻĄÉņŚÉņä£ SMoVņØś 3ŌĆ▓ UTRņØĆ RNA1Ļ│╝ RNA2 Ļ░äņŚÉ Ēü░ ņ░©ņØ┤Ļ░Ć ņŚåļŖö Ļ▓āņØä ĒÖĢņØĖĒĢśņśĆļŗż. ĒĢ£ĒÄĖ, 3ŌĆ▓ ļ¦Éļŗ©ņŚÉ poly A tailņØä Ļ░Ćņ¦ĆļŖö ļ░öņØ┤ļ¤¼ņŖżņØś ĒŖ╣ņ¦ĢņØĖ polyadenylation signal (AATAAA motif)ņØ┤ 3ŌĆ▓ ļ¦Éļŗ©ņØś ņāüļŗ© 174ņŚ╝ĻĖ░ņŚÉ ņ£äņ╣śĒĢśļŖö Ļ▓āņØä ĒÖĢņØĖĒĢśņśĆļŗż(Fig. 1). SMoVļŖö ņĀäņäĖĻ│äņĀüņ£╝ļĪ£ ļöĖĻĖ░(Fragaria spp.)ņŚÉņä£ ļŗżņ¢æĒĢ£ ļČäļ”¼ņŻ╝ļōżņØ┤ ļ│┤Ļ│ĀļÉśņ¢┤ ņ׳ļŖö ļ░śļ®┤(Bhagwat ļō▒, 2016; ThompsonĻ│╝ Jelkmann, 2003; Thompson ļō▒, 2002; Yang ļō▒, 2007) ĻĄŁļé┤ ļČäļ”¼ņŻ╝ņŚÉ ļīĆĒĢ£ ņŚ╝ĻĖ░ņä£ņŚ┤ ļō▒ļĪØņØĆ ļ│Ė ņŚ░ĻĄ¼ļź╝ ĒåĄĒĢ┤ ņ▓śņØī ļ│┤Ļ│ĀļÉśņŚłļŗż. ĻĄŁļé┤ ļČäļ”¼ņŻ╝ņØś ļČäņ×ÉĻ│äĒåĄĒĢÖņĀü ņ£äņ╣śļź╝ ļČäņäØĒĢśĻĖ░ ņ£äĒĢ┤ GenBankņŚÉ ļō▒ļĪØļÉ£ ļŗżņ¢æĒĢ£ SMoV ļČäļ”¼ņŻ╝(Table 3)ņØś 3ŌĆ▓ UTR ņŚ╝ĻĖ░ņä£ņŚ┤ņØä ņłśņ¦æĒĢśņŚ¼ maximum likelihood ļČäņäØ ļ░®ņŗØņØä ņØ┤ņÜ®ĒĢ£ Ļ│äĒåĄļČäņäØņØä ņŗżņŗ£ĒĢśņśĆļŗż. Phylogenetic tree ņ×æņä▒ Ļ▓░Ļ│╝, ĻĄŁļé┤ ļČäļ”¼ņŻ╝ ļīĆļČĆļČäņØĆ ņ║Éļéśļŗż ļČäļ”¼ņŻ╝ņÖĆ ĻĘ╝ņŚ░Ļ┤ĆĻ│äĻ░Ć Ļ░Ćņן Ļ░ĆĻ╣īņÜ┤ ļ░śļ®┤ SH-PņØś Ļ▓ĮņÜ░ ņ£Āļ¤Į ļ░Å ņØ┤ļ×Ć ļČäļ”¼ņŻ╝ņÖĆ ĻĘ╝ņŚ░Ļ┤ĆĻ│äļź╝ ĒśĢņä▒ĒĢśļŖö Ļ▓āņØä ņĢī ņłś ņ׳ņŚłļŗż. ļśÉĒĢ£ ņŚ╝ĻĖ░ņä£ņŚ┤ņØś ņ¦äĒÖöņĀü ņĖĪļ®┤ņŚÉņä£ SMoVņØś ļ░£ņāØņØ┤ ļ¦ÄņØĆ ņ£Āļ¤ĮĻ│╝ ņ║Éļéśļŗż ļČäļ”¼ņŻ╝ņØś Ļ▓ĮņÜ░ņŚÉļŖö ĻĄŁļé┤ ļČäļ”¼ņŻ╝ņÖĆ ļ╣äĻĄÉĒ¢łņØä ļĢī ņŚ╝ĻĖ░ņä£ņŚ┤ņ¦äĒÖöņØś ļČäĒÖöĻ░Ć ļŹö ļ¦ÄņØ┤ ņ¦äĒ¢ēļÉ£ Ļ▓āņŚÉ ļ╣äĒĢ┤ ĻĄŁļé┤ ļČäļ”¼ņŻ╝ļŖö ļ¬©ļæÉ ņ¦äĒÖöņĀüņ£╝ļĪ£ ļČäĒÖöĻ░Ć ļÉśņ¦Ć ņĢŖņĢśņØīņØä ĒÖĢņØĖĒĢĀ ņłś ņ׳ņŚłļŗż(Fig. 2). ņØ┤ļ¤¼ĒĢ£ Ļ▓░Ļ│╝ļĪ£ ļ»ĖļŻ©ņ¢┤ ļ│╝ ļĢī ĻĄŁļé┤ņŚÉņä£ļŖö SMoV ļ░£ņāØņØ┤ ļ¦Äņ¦Ć ņĢŖņĢä ļ░öņØ┤ļ¤¼ņŖżņØś ļ│ĆņØ┤Ļ░Ć ņØ╝ņ¢┤ļéĀ ĒÖĢļźĀņØ┤ ņĀüĻ│Ā Ļ░ÉņŚ╝ļÉ£ ļ¬©ņŻ╝ļĪ£ļČĆĒä░ ņśüņ¢æļ▓łņŗØņŚÉ ņØśĒĢ┤ Ļ▓ĆņČ£ļÉśĻ│Ā ņ׳ļŖö Ļ▓āņ£╝ļĪ£ ņČöņĖĪļÉ£ļŗż.
Table┬Ā3
Information of SMoV isolates analyzed in this study
| Isolate | Hosta | Year | Country | Accession No. | Reference |
|---|---|---|---|---|---|
| MH | F. ├Ś ananassa cv. Mihong | 2017 | Korea | MG418837 | Present study |
| MaeH | F. ├Ś ananassa cv. Maehyang | 2018 | Korea | MN520311 | Present study |
| SH-P | F. ├Ś ananassa cv. Seolhyang | 2018 | Korea | MN520312 | Present study |
| ST | F. ├Ś ananassa cv. Santa | 2018 | Korea | MN520314 | Present study |
| GS-G | F. ├Ś ananassa cv. Geumsil | 2018 | Korea | MN520309 | Present study |
| GS-S | F. ├Ś ananassa cv. Geumsil | 2019 | Korea | MN520310 | Present study |
| SH-S | F. ├Ś ananassa cv. Seolhyang | 2019 | Korea | MN520313 | Present study |
| 1134 | F. vesca | 1987 | Netherland | AJ311876 | Thompson et al. (2002) |
| 197-3A | F. ├Ś ananassa | 1989 | Czech Republic | AJ496589 | Thompson and Jelkmann (2003) |
| 197-3B | F. vesca UC4 | 1989 | Czech Republic | AJ496590 | Thompson and Jelkmann (2003) |
| 1141 | F. vesca UC5 | 1987 | Netherland | AJ496581 | Thompson and Jelkmann (2003) |
| 1166 | F. vesca UC5 | 1987 | Netherland | AJ496582 | Thompson and Jelkmann (2003) |
| 1265 | F. vesca UC5 | 1987 | Netherland | AJ496584 | Thompson and Jelkmann (2003) |
| 1266 | F. vesca UC5 | 1987 | Netherland | AJ496585 | Thompson and Jelkmann (2003) |
| 1278 | F. vesca UC5 | 1987 | Netherland | AJ496586 | Thompson and Jelkmann (2003) |
| 1280 | F. vesca UC5 | 1987 | Netherland | AJ496587 | Thompson and Jelkmann (2003) |
| 1509-4 | F. vesca UC5 | 1999 | Poland | AJ496588 | Thompson and Jelkmann (2003) |
| 1248 | F. virginiana | 1999 | Poland | AJ496583 | Thompson and Jelkmann (2003) |
| 3CH | F. chiloensis | 2001 | Chile | AJ496591 | Thompson and Jelkmann (2003) |
| CN1 | F. ├Ś ananassa cv. Hokowase | 2005 | China | AY919307 | Yang et al. (2007) |
| NSper3 | F. ├Ś ananassa | 2013 | Canada | KU200457 | Bhagwat et al. (2016) |
| NSper17 | F. ├Ś ananassa | 2013 | Canada | KU200459 | Bhagwat et al. (2016) |
| NSper51 | F. ├Ś ananassa | 2013 | Canada | KU200461 | Bhagwat et al. (2016) |
| NB926 | F. ├Ś ananassa | 2014 | Canada | KU200454 | Bhagwat et al. (2016) |
| Ontario | F. ├Ś ananassa | 2014 | Canada | KU177219 | Bhagwat et al. (2016) |
| AG | F. ├Ś ananassa | 2018 | Iran | MK303324 | Unpublished |
| K3 | F. ├Ś ananassa | 2018 | Iran | MK303320 | Unpublished |
| K9 | F. ├Ś ananassa | 2018 | Iran | MK303322 | Unpublished |
| SQA | F. ├Ś ananassa | 2018 | Iran | MK303323 | Unpublished |
| SS2 | F. ├Ś ananassa | 2018 | Iran | MK303321 | Unpublished |
Fig.┬Ā1
An alignment of nucleotide sequences of the 3ŌĆ▓ untranslated region of 7 Korean isolates of Strawberry mottle virus and RNA1 and RNA2 of the isolate 1134. Sequences were aligned by ClustalX2. The consensus sequences are shown at the top. For each position, bases in a majority and bases in a minority are showed in the light gray background and white background, respectively. Extra bases only found in the Korean isolates are boxed. Polyadenylation signal (AATAAA motif at position 164-169), is underlined. Accession numbers of the isolates in Table 3.
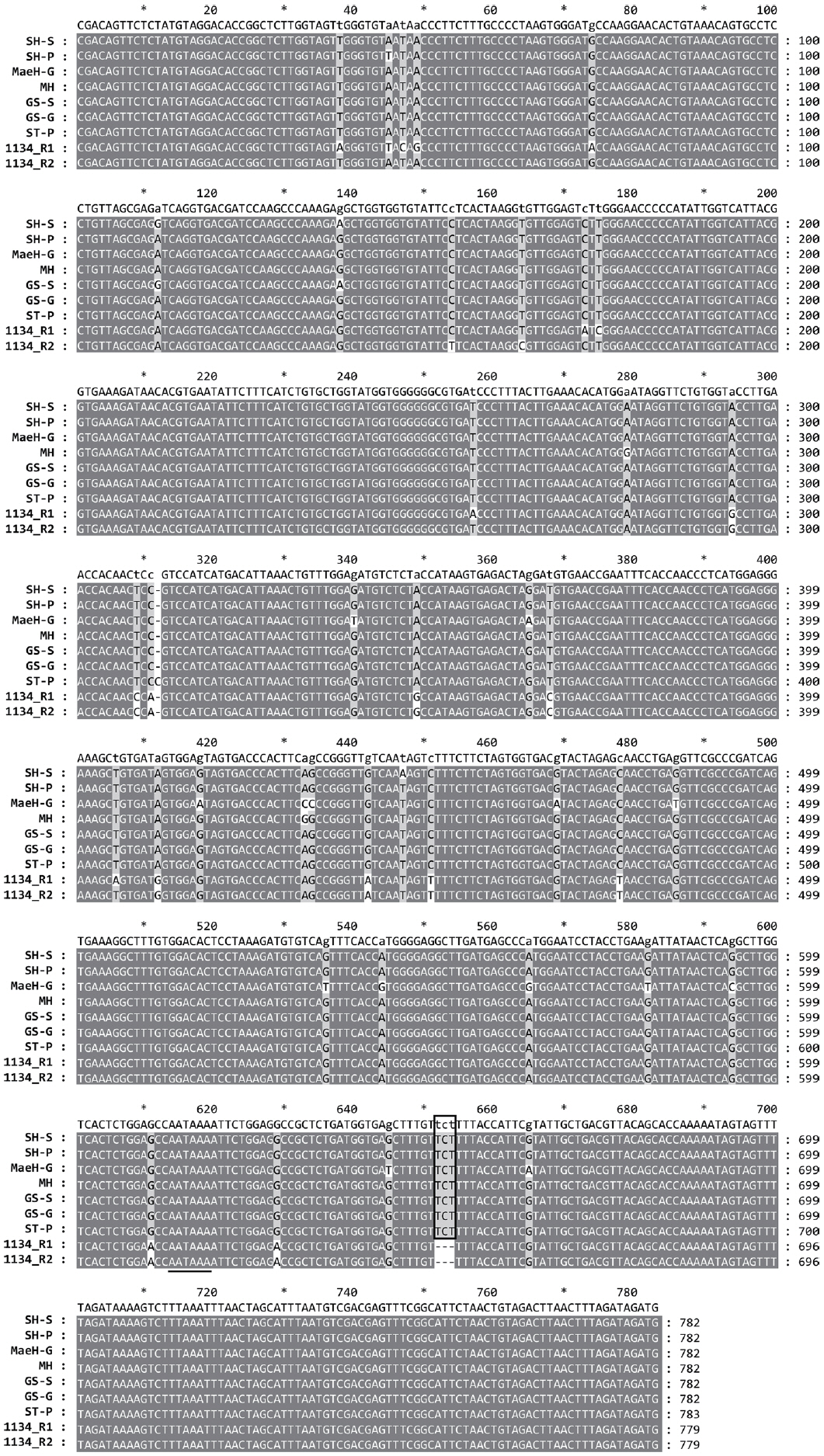
Fig.┬Ā2
Phylogenetic analysis of the 3ŌĆ▓ untranslated region sequences of Strawberry mottle virus isolates. The tree was constructed by the maximum likelihood method using MEGA 7 program. Seven Korean isolates are marked with arrows. The numbers on the branches indicate bootstrap percentages based on 1,000 replications (only values >60% are shown). Description and accession numbers of the isolates are shown in Table 3. The Black raspberry necrosis virus (BRNV, GenBank accession No. NC008183) was included as an outgroup control.

ĻĄŁļé┤ ļöĖĻĖ░ ļåŹĻ░ĆņØś ņ¦äļö¦ļ¼╝ ļ░£ņāØ ņóģ ļÅÖņĀĢ
ĻĄŁļé┤ ļöĖĻĖ░ņØś SMoV Ļ░ÉņŚ╝Ļ│╝ ņ¦äļö¦ļ¼╝ ņĀäņŚ╝ Ļ┤ĆĻ│äļź╝ ĒīīņĢģĒĢśĻĖ░ ņ£äĒĢ┤ ĻĄŁļé┤ ļöĖĻĖ░ ļåŹĻ░ĆņØś ņŻ╝ņÜö ļ░£ņāØ ņ¦äļö¦ļ¼╝ ņóģņØä ļÅÖņĀĢĒĢśņśĆļŗż. ņł£ņ░ĮĻ│╝ ņ¦äņŻ╝, ĒĢ©ņ¢æ ļöĖĻĖ░ļåŹĻ░ĆņŚÉņä£ ļåŹĻ░Ćļ│äļĪ£ 20ļ¦łļ”¼ņö® ņ¦äļö¦ļ¼╝ņØä ņ▒äņ¦æĒĢśņŚ¼ DNA barcoding ļ░®ļ▓Ģ(Lee ļō▒, 2011)ņŚÉ ļö░ļØ╝ cytochrome c oxidase subunit IņØś ņŚ╝ĻĖ░ņä£ņŚ┤ņØä ļČäņäØĒĢ£ Ļ▓░Ļ│╝, 84%Ļ░Ć ļ¬®ĒÖöņ¦äļö¦ļ¼╝ļĪ£ ļéśĒāĆļé¼Ļ│Ā 16%ļŖö Aphis cf. rostellaĻ░Ć ļ░£ņāØĒĢśļŖö Ļ▓āņ£╝ļĪ£ ņĪ░ņé¼ļÉśņŚłļŗż(data not shown). ņŻ╝ņÜö ļöĖĻĖ░ņ¦äļö¦ļ¼╝ļĪ£ ņĢīļĀżņ¦ä Chaetosiphon spp.ļŖö ļ░£Ļ▓¼ļÉśņ¦Ć ņĢŖņĢśļŗż. ļöĖĻĖ░ ņ×¼ļ░░ ĒŖ╣ņä▒ņāü ļåŹĻ░Ćļ│äļĪ£ ņĀĢĻĖ░ņĀüņØĖ ļ░®ņĀ£ ņ▓śļ”¼ļĪ£ ņØĖĒĢ┤ ļ¦ÄņØĆ ņłśņØś ņ¦äļö¦ļ¼╝ņØä ņ▒äņ¦æĒĢśĻĖ░ņŚÉļŖö ņĀ£ĒĢ£ņØ┤ ņ׳ņŚłļŗżļŖö ņĀÉņØä Ļ░ÉņĢłĒĢśņŚ¼ļÅä ĻĄŁļé┤ņŚÉņä£ ļöĖĻĖ░ ļ░öņØ┤ļ¤¼ņŖżņØś ņŻ╝ņÜö ņ¦äļö¦ļ¼╝(Chaetosiphon spp.)ņØĆ Ēü¼Ļ▓ī ļ¼ĖņĀ£Ļ░Ć ļÉśņ¦Ć ņĢŖļŖö Ļ▓āņ£╝ļĪ£ ļ│┤ņŚ¼ņ¦Ćņ¦Ćļ¦ī SMoVņØś Ļ▓ĮņÜ░ ļ¬®ĒÖöņ¦äļö¦ļ¼╝ļĪ£ļÅä ņĀäļ░śņØ┤ ļÉśĻĖ░ ļĢīļ¼ĖņŚÉ ņŻ╝ņØśĻ░Ć ĒĢäņÜöĒĢ┤ ļ│┤ņØĖļŗż.
ļ│Ė ņŚ░ĻĄ¼ļź╝ ĒåĄĒĢ┤, ĻĄŁļé┤ņŚÉņä£ ņ×¼ļ░░ļÉśļŖö ņŻ╝ņÜö ļöĖĻĖ░ ĒÆłņóģņØś ņ¦äļö¦ļ¼╝ ņĀäļ░ś ļ░öņØ┤ļ¤¼ņŖż Ļ░ÉņŚ╝ļźĀņØĆ 1% ļé┤ņÖĖ(SMYEV, 0.7%; SMoV, 1.3%)ļĪ£ ņĪ░ņé¼ļÉśņŚłĻ│Ā, ļ░öņØ┤ļ¤¼ņŖż Ļ░ÉņŚ╝ņŚÉ ļö░ļźĖ ņāØņ£Ī ņĀĆĒĢśļéś ļÜ£ļĀĘĒĢ£ ļ│æņ¦ĢņØĆ Ļ┤Ćņ░░ļÉśņ¦Ć ņĢŖņĢśļŗż. SMoV ĻĄŁļé┤ ļČäļ”¼ņŻ╝ 7Ļ░£ņŚÉ ļīĆĒĢ£ ņŚ╝ĻĖ░ņä£ņŚ┤ ļ░Å ņ£ĀņĀäĒĢÖņĀü Ļ│äĒåĄļČäņäØņØä ĒåĄĒĢ┤ ĻĄŁļé┤ SMoV ļČäļ”¼ņŻ╝ļŖö ņ¦äĒÖöņĀüņ£╝ļĪ£ ļČäĒÖöĻ░Ć ļ¦Äņ¦Ć ņĢŖņØīņØä ĒÖĢņØĖĒĢśņśĆļŗż. ņ¦äļö¦ļ¼╝ ņóģ ļČäņäØ Ļ▓░Ļ│╝ ņŻ╝ņÜö ļöĖĻĖ░ ņ¦äļö¦ļ¼╝(Chaetosiphon spp.)ņØś ļ░£ņāØļÅä ņŚåļŖö Ļ▓āņ£╝ļĪ£ ņĪ░ņé¼ļÉśņ¢┤ ņ¦äļö¦ļ¼╝ ņĀäļ░śņŚÉ ņØśĒĢ£ Ļ░ÉņŚ╝ļ│┤ļŗżļŖö ņśüņ¢æļ▓łņŗØņŚÉ ņØśĒĢ┤ ņĀäļ░śļÉśņ¢┤ ļ░£ņāØļÉśĻ│Ā ņ׳ņØä ĒÖĢļźĀņØ┤ ļåÆņØä Ļ▓āņ£╝ļĪ£ ņČöņĖĪļÉ£ļŗż. ĻĄŁļé┤ņØś Ļ▓ĮņÜ░, ļöĖĻĖ░ ĒÆłņóģ ņ£ĪņóģņŚÉ ņ׳ņ¢┤ņä£ ļ░öņØ┤ļ¤¼ņŖż ļ¼┤ļ│æļ¼ś ņāØņé░ņ▓┤Ļ│äĻ░Ć ĒÜ©ņ£©ņĀüņ£╝ļĪ£ ņ¦äĒ¢ēļÉśĻ│Ā ņ׳ļŗżĻ│Ā ņŚ¼Ļ▓©ņ¦Ćļ®░, ļåŹĻ░ĆņŚÉ ļ│┤ĻĖē ĒøäņŚÉļÅä ņ×¼ļ░░Ļ┤Ćļ”¼Ļ░Ć ņל ļÉśĻ│Ā ņ׳ļŖö Ļ▓āņ£╝ļĪ£ ļ│┤ņŚ¼ņ¦äļŗż. ļŹö ļéśņĢäĻ░Ć ņāłļĪ£ņÜ┤ ļöĖĻĖ░ ļ░öņØ┤ļ¤¼ņŖż ļ░Å ļ¦żĻ░£ņČ® ņóģ ņ£Āņ×ģņŚÉ ļīĆļ╣äĒĢśĻ│Ā ĻĖ░ņĪ┤ņŚÉ ļ│┤Ļ│ĀļÉ£ ļ░öņØ┤ļ¤¼ņŖżņŚÉ ļīĆĒĢ┤ņä£ļŖö ļöĖĻĖ░ ĒÆłņóģļ│äļĪ£ ļ│æņøÉņä▒ ļČäņäØņŚÉ ļīĆĒĢ£ ņŚ░ĻĄ¼Ļ░Ć ļŹö ņ¦äĒ¢ēļÉśņ¢┤ņĢ╝ ĒĢĀ Ļ▓āņ£╝ļĪ£ ņāØĻ░üļÉ£ļŗż.
ņÜö ņĢĮ
ļöĖĻĖ░ļź╝ Ļ░ÉņŚ╝ĒĢśļŖö 30ņŚ¼ ņóģņØś ļ░öņØ┤ļ¤¼ņŖż ņżæ ņĀä ņäĖĻ│äņĀüņ£╝ļĪ£ Ļ░Ćņן ļ¦ÄņØ┤ ļ░£ņāØĒĢśļŖö 4ņóģņØś ļ░öņØ┤ļ¤¼ņŖżļŖö Strawberry mild yellow edge virus (SMYEV), Strawberry mottle virus (SMoV), Strawberry crinkle virus (SCV), Strawberry vein banding virus (SVBV)ļĪ£ ņØ┤ļōżņØĆ ļ¬©ļæÉ ņ¦äļö¦ļ¼╝ņØ┤ ņĀäļ░śļÉśļ®░ Ļ▓ĮņĀ£ņĀüņ£╝ļĪ£ Ļ░Ćņן ņżæņÜöĒĢ£ ļ░öņØ┤ļ¤¼ņŖżļōżņØ┤ļŗż. 2018-2019ļģäĻ╣īņ¦Ć ĻĄŁļé┤ ņŻ╝ņÜö ļöĖĻĖ░ ņāØņé░ņ¦ĆņŚÉņä£ ĻĄŁļé┤ ļöĖĻĖ░ ĒÆłņóģņØä ļīĆņāüņ£╝ļĪ£ ņØ┤ļōż 4ņóģ ņ¦äļö¦ļ¼╝ ņĀäļ░ś ļ░öņØ┤ļ¤¼ņŖżņØś ļ░£ņāØņĪ░ņé¼ļź╝ ņŗżņŗ£ĒĢśņśĆļŗż. ĻĘĖ Ļ▓░Ļ│╝, ņØ╝ļČĆ ĻĄŁļé┤ ļöĖĻĖ░ ĒÆłņóģņŚÉņä£ SMYEVņÖĆ SMoVĻ░Ć Ļ░üĻ░ü 0.7%ņÖĆ 1.3%ņØś ļé«ņØĆ Ļ░ÉņŚ╝ļźĀļĪ£ Ļ▓ĆņČ£ļÉśņŚłņ£╝ļ®░ SCVņÖĆ SVBVļŖö ņĀäĒśĆ Ļ▓ĆņČ£ļÉśņ¦Ć ņĢŖņĢśļŗż. ĒĢ£ĒÄĖ, ļ░öņØ┤ļ¤¼ņŖż Ļ░ÉņŚ╝ ņŗØļ¼╝ņŚÉņä£ ļ│æņ¦ĢņØĆ Ļ┤Ćņ░░ļÉśņ¦Ć ņĢŖņĢśļŗż. ĻĄŁļé┤ņŚÉņä£ SMoVņŚÉ ļīĆĒĢ£ ņŚ╝ĻĖ░ņä£ņŚ┤ņØĆ ļ│┤Ļ│ĀļÉ£ ļ░ö ņŚåņ¢┤ SMoV ĻĄŁļé┤ļČäļ”¼ņŻ╝ņŚÉ ļīĆĒĢ£ 3ŌĆ▓ untranslated regionņØś ņŚ╝ĻĖ░ņä£ņŚ┤ņØä Ļ▓░ņĀĢĒĢśĻ│Ā ļČäņäØĒĢśņśĆļŗż. ĻĖ░ņĪ┤ņŚÉ ļ│┤Ļ│ĀļÉ£ SMoV ļČäļ”¼ņŻ╝ļōżĻ│╝ņØś ļČäņ×ÉĻ│äĒåĄĒĢÖņĀü ņ£ĀņŚ░Ļ┤ĆĻ│ä ļČäņäØ Ļ▓░Ļ│╝, ļīĆļČĆļČäņØś ĻĄŁļé┤ ļČäļ”¼ņŻ╝ļŖö ņ║Éļéśļŗż ļČäļ”¼ņŻ╝ņÖĆ ĻĘ╝ņŚ░Ļ┤ĆĻ│äĻ░Ć ļåÆņØĆ Ļ▓āņ£╝ļĪ£ ļéśĒāĆļé¼ņ£╝ļ®░ ņŚ╝ĻĖ░ņä£ņŚ┤ņØś ņ¦äĒÖöņĀü ņĖĪļ®┤ņŚÉņä£ ļČäĒÖöĻ░Ć Ļ▒░ņØś ņØ╝ņ¢┤ļéśņ¦Ć ņĢŖņØĆ ļČäļ”¼ņŻ╝ņ×äņØä ĒÖĢņØĖĒĢśņśĆļŗż.
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






