바이러스 열성 저항성: 병저항성 작물 개발을 위한 유전자 교정 소재 발굴 연구의 동향
Recessive Resistance: Developing Targets for Genome Editing to Engineer Viral Disease Resistant Crops
Article information

Abstract
Plant viruses are among the important pathogens that cause severe crop losses. The most efficient method to control viral diseases is currently to use virus resistant crops. In order to develop the virus resistant crops, a detailed understanding of the molecular interactions between viral and host proteins is necessary. Recessive resistance to a pathogen can be conferred when plant genes essential in the life cycle of a pathogens are deficient, while dominant resistance is mediated by host resistance (R) genes specifically interacting with effector proteins of pathogens. Thus, recessive resistance usually works more stably and broadly than dominant resistance. While most of the recessive resistance genes have so far been identified by forward genetic approaches, recent advances in genome editing technologies including CRISPR/Cas9 have increased interest in using these technologies as reverse genetic tools to engineer plant genes to confer recessive resistance. This review summarizes currently identified recessive resistance genes and introduces reverse genetic approaches to identify host interacting partner proteins of viral proteins and to evaluate the identified genes as genetic resources of recessive resistance. We further discuss recent advances in various precise genome editing technologies and how to apply these technologies to engineer plant immunity.
서 론
식물 바이러스는 살아 있는 세포에서만 증식하는 절대기생체로, 매우 단순한 유전체를 가지고 있다. 바이러스의 유전체는 단일 가닥 또는 이중 가닥의 RNA 또는 DNA로 이루어져 있으며 바이러스의 생활사에 꼭 필요한 단백질 정보를 포함하고 있다. 식물 바이러스병에 대한 치료 약제가 개발되어 있지 않고, 바이러스 복제 중 돌연변이가 쉽게 발생하기 때문에 작물 생산량 손실을 일으키는 주요 요인으로 잘 알려져 있다. 뿐만 아니라, 세계적으로 나타나고 있는 기후 변화와 국제화로 인한 무역 다변화로 인하여 식물 바이러스 질병 유입 및 발생의 위험이 크게 증가하고 있다(Hashimoto 등, 2016a).
작물의 안정적 생산을 위해 식물 바이러스 질병을 방제하기 위한 다양한 노력이 이루어지고 있다. 전통적인 식물 바이러스 방제 전략은 대부분 살충제 또는 천적을 이용해 매개충을 제거하는 것에 초점이 맞추어져 있었다(Legg 등, 2014). 그러나, 바이러스 질병의 발생과 관련된 매개충의 이동 역학, 빠른 바이러스의 진화, 예상 불가능한 바이러스의 기주 범위 확장 등 복잡한 역학적 요소들로 인해 장기적인 방제 전략을 세우는 것은 매우 어렵다(Loebenstein 와 Katis, 2014). 이에 따라, 현재로서 바이러스를 방제하기 위한 가장 효과적인 방법은 바이러스에 저항성을 가진 품종을 재배하는 것이다.
식물에서의 선천적 병저항성은 크게 두 가지 양상으로 발현된다 (Fig. 1). 하나는 저항성 유전자(resistance gene, R gene)에 의해 매개되는 우성 저항성(dominant resistance)이고, 다른 하나는 병원체의 감염에 필수적인 기주 인자의 결여에 의해 나타나는 열성 저항성(recessive resistance)이다(Hashimoto 등, 2016a). 우성 저항성을 매개하는 R 유전자는 전통적인 교배 육종을 통해 병저항성 품종을 개발하는데 널리 이용되어 왔으며, 병원체에서 유래한 비병원성 유전자(avirulence gene, Avr gene)와의 상호작용을 통해 저항성을 유도한다(Keen, 1990). 우성 저항성은 특이적 반응을 통해 대상 병원체에 대해 강한 저항성을 유발하지만, R-Avr 상호작용의 특이성에 따라 병원체 인식의 스펙트럼이 좁고, 돌연변이가 빠르게 발생하는 바이러스의 경우에 쉽게 저항성이 무너질 수 있다(Langner 등, 2018). 이에 반해, 열성 저항성은 병원체가 생존을 위해 필요로 하는 기주 유전자가 결여되거나 상호작용이 억제되었을 때 나타난다. 실제로, 지금까지 작물에서 알려진 바이러스 저항성과 관련된 유전자의 절반은 열성 저항성 유전자이다(Kang 등, 2005). 일반적으로, 열성 저항성은 우성 저항성보다 강하게 작용하며 다소 넓은 특이성으로 돌연변이 발생에 의해 저항성이 쉽게 무너지지 않는 특징이 있다(Hashimoto 등, 2016a; Truniger 와 Aranda, 2009; Wang과 Krishnaswamy, 2012).

Schematic representation of dominant and recessive resistance in a plant-virus pathosystem. A virus-encoded protein can act as an avirulence factor that is recognized by a host-encoded resistance (R) protein. The interaction between a viral avirulence factor and a host R protein triggers down-stream defense responses, resulting in activation of dominant resistance. Plant viruses are obligate parasites and require various host-encoded proteins (host factors) to complete the steps of their life cycle. Therefore, the absence of appropriate host factors or inhibition of the interactions between viral proteins and corresponding host factors may confer recessive resistance.
지금까지 밝혀진 대부분의 바이러스에 대한 열성 저항성 유전자는 진핵 생물의 번역 개시 인자(translation initiation factors)인 eIF4E와 eIF4G 또는 이들의 아형(isoform)들이다 (Robaglia 와 Caranta, 2006) (Table 1). eIF4E 돌연변이 식물에서 실제로 포티바이러스와 다른 몇몇의 바이러스에 대한 감수성(susceptibility)이 상실되는 것이 확인된 바 있다(Hashimoto 등, 2016a; Robaglia와 Caranta, 2006). 이외의 다양한 바이러스에 대한 열성 저항성을 유도하기 위해서는 바이러스의 증식에 필수적인 다양한 기주 인자를 밝혀내고 이들의 열성 저항성 유전 자원 활용성을 확보하는 것이 중요하다. 이러한 기초 연구 결과를 바탕으로 유전자 편집 기술을 활용하여 기주 유전자에 대한 정교한 돌연변이를 도입하는 것으로 열성 저항성을 유도할 수 있기 때문이다.

Antiviral recessive resistance genes associated with translation initiation
본 논문에서는 기존에 밝혀진 바이러스병에 대한 열성 저항성 유전자에 대해 소개하는 한편, 새로운 열성 저항성 유전 자원 발굴을 위한 최근 연구 동향 및 유전자 편집 기술 적용을 통한 열성 저항성 작물 개발 연구의 중요성을 정리하였다.
기존에 밝혀진 열성 저항성 유전자
열성 저항성은 바이러스와 기주 식물 간의 분자적 상호작용을 기반으로 이루어진다. 식물 바이러스는 다양한 기주 세포의 단백질을 이용하여 자신들의 유전체를 증식 시킨 다음, 건강한 세포나 조직으로 이동한다. 바이러스 생활사에 중요한 식물 유전자에 돌연변이가 발생하여 이용이 억제되면 식물은 바이러스에 대한 열성 저항성을 획득할 수 있다(Fraser, 1990). 지금까지 발견된 약 200여개 이상의 바이러스 저항성 유전자 중에서 약 절반에 해당하는 저항성 유전자가 열성 저항성과 관련된 유전자이다(Kang 등, 2005). 열성 저항성 유전자는 병감수성 상실(loss-of-susceptibility)이 나타난 돌연변이 식물을 이용하여 찾거나 자연적으로 진화한 생태형(ecotype)에서 저항성이 유도된 것들을 찾는 과정을 통해 선발되었다(Diaz-Pendon 등, 2004; Truniger 와 Aranda, 2009) (Table 2). 놀랍게도, 현재까지 밝혀진 열성 저항성 유전자를 분석한 결과 대부분 번역 개시 인자(translation initiation factor)인 eIF4E와 eIF4G, 그리고 그의 아형(isoform)들인 것으로 확인되었다(Robaglia 와 Caranta, 2006). 그러나 자연적으로 형성되는 열성 저항성 유전자에 대한 연구는 아직까지 부족한 상황이며, 돌연변이 선발 연구를 통하여 eIF4E와 eIF4G 이외에도 다양한 감수성 결정 인자가 존재한다는 것이 확인되었기 때문에 이와 관련된 체계적인 연구가 필요하다.

The genetic resources for recessive resistance found in loss-of-susceptibility mutants and naturally occurring resistant cultivars
열성 저항성은 다른 바이러스보다 포티바이러스 그룹에 속하는 바이러스들에서 많이 연구되었으며, 지금까지 밝혀진 열성 저항성 유전자 목록에서도 절반 이상이 포티바이러스에 해당된다(Table 1). 포티바이러스 그룹이 아닌 다른 바이러스에서도 eIF4E 매개 저항성이 나타나는 것으로 관찰되었는데, eIF4E 매개 열성 저항성은 최소 한가지 이상의 eIF4E와 상호작용을 하는 바이러스들에 대해서만 효과적인 것으로 확인되었다(Hashimoto 등, 2016a). 그러나, 기능적으로 다중성(redundancy)이 있는 eIF4E들이 있기 때문에 한 가지의 eIF4E의 결함이 항상 식물 바이러스에 대한 저항성유도로 이어지지는 않을 수 있다(Martinez-Silva 등, 2012; Mayberry 등, 2011). 그리고 식물이 살아가기 위해 꼭 필요한 eIF4E의 기능이 있기 때문에 eIF4E knockout 돌연변이체에서 배아 치사(embryo-lethal) 표현형이 나타나기도 한다(Nicaise 등, 2007; Patrick 등, 2014). 이처럼, eIF4E 또는 아형만을 이용하여 열성 저항성을 유도하는 것은 한계가 있기 때문에 더 다양한 바이러스 종에 대응할 수 있는 추가적인 열성 저항성 유전자 소재를 찾는 연구가 필요하다.
eIF4E에 의해 매개되는 포티바이러스에 대한 열성 저항성
eIF4E은 번역 개시 복합체를 이루는 구성 요소 중 하나로, 다른 단백질들을 복합체로 이동시키는 기능과 함께 mRNA의 캡 결합 단백질(cap binding protein)의 기능을 한다(Gazo 등, 2004). eIF4E의 구조와 기능은 모든 진핵 생물 사이에서 매우 잘 보존되어 있으며, 바이러스 RNA와 직접적으로 상호작용하거나 바이러스 단백질을 통해 바이러스 RNA에 결합하기도 한다. 포티바이러스는 viral protein genome linked (VPg) 단백질을 통해 직접적으로 eIF4E와 상호작용을 한다(Roudet-Tavert 2007). VPg는 다른 바이러스 단백질이나 기주 단백질과 상호작용하며 바이러스의 복제, 이동과 관련된 많은 과정들을 조절하는 허브 단백질로 작용을 한다고 알려져 있다(Jiang과 Laliberte, 2011). VPg의 전구체 형태나 기능에 따라 세포내 축적 위치가 다양한 것이 특징이다.
이러한 포티바이러스의 VPg와 식물의 eIF4E간의 상호작용은 열성 저항성과 크게 관련이 있다(Lellis 등, 2002; Wang과 Krishnaswamy, 2012). 포티바이러스에 대한 열성 저항성은 자연적으로 발생한 eIF4E 돌연변이가 VPg와 상호작용하는 능력을 잃었을 때 바이러스 유전자의 번역이 억제되면서 유도된다. 하지만 이러한 돌연변이가 기주의 mRNA 번역은 방해하지 않는다. 다양한 eIF4E 저항성 유전자는 애기장대에서 바이러스에 대한 감수성 상실(loss-of-susceptibility)이 발생한 돌연변이체 선발이나 자연적으로 발생한 열성 저항성 유전자를 찾는 과정에서 발견이 되었고, 다른 작물에서 실제로 저항성이 유도되는지 연구를 통해 확인되었다(Lellis 등, 2002; Ruffel 등, 2002; Yoshii 등, 2004). eIF4E 매개 열성 저항성은 포티바이러스에 속하는 tobacco etch virus에 대한 감수성 상실(loss-of-susceptibility)이 나타난 eIF(iso)4E 애기장대 돌연변이체에서 처음으로 발견되었다(Lellis 등, 2002). 이후의 연구에서 포티바이러스에 대한 eIF4E매개 저항성이 고추(Capsicum annuum), 상추(Lactuca sativa), 야생 토마토(Solanum habrochaites) 등 몇몇의 저항성 작물 품종에서 나타나는 것을 확인했다(Nicaise 등, 2003; Ruffel 등, 2005). 최근에는 애기장대에서 최신 유전자 교정 기술인 CRISPR/Cas9을 이용해 eIF4E 유전자에 직접적으로 돌연변이를 일으켜 포티바이러스에 대한 열성 저항성이 유도된 돌연변이 식물을 제작하기도 했다(Pyott 등, 2016). 이렇게 열성 저항성을 획득한 애기장대는 야생형(wild type)과 비교했을 때 생장 등 다른 표현형적인 차이가 나타나지 않았다(Pyott 등, 2016).
열성 저항성 획득을 위한 후보 소재인 기주 인자 연구 동향
열성 저항성은 바이러스가 필요로 하는 기주 인자 혹은 이들과의 상호작용이 결여되면서 획득된다. 따라서, 바이러스가 증식하는데 필요로 하는 기주 인자, 특히 바이러스 단백질과 직접적으로 상호작용하는 기주 단백질을 밝히는 것은 열성 저항성 유전 소재 탐색에 있어 중요한 접근법이라 할 수 있다.
식물이 바이러스에 감염되는 과정은 바이러스 입자의 분해, 바이러스 게놈의 번역, 바이러스 증식 복합체(virus replication complex, VRC) 형성 및 게놈 복제, 바이러스 입자 형성(genome encapsidation), 세포 간 이동(cell-to-cell movement), 장거리 수송(long-distance transport) 등 여러 가지 단계로 나누어지는 매우 복잡한 과정으로, 다양한 기주 인자들이 바이러스 감염 과정의 각 단계에서 밀접하게 작용하는 것으로 밝혀지고 있다 (Ahlquist 등, 2003; Nagy 와 Richardson, 2012; Wang, 2015).
주로 애기장대나 일부 모델 식물 또는 효모를 이용해서 순행 유전학(forward genetics) 또는 역유전학(reverse genetics) 접근 방식으로 바이러스 증식에 필수적인 기주 인자를 밝히려는 연구들이 많이 수행되었다(Hashimoto 등, 2016a; Wang과 Krishnaswamy, 2012). 특히 바이러스 단백질과 직접적으로 상호작용하는 기주 인자 또는 바이러스 단백질을 포함하는 단백질 복합체의 구성 요소로 작용하는 기주 인자를 알아내는 연구가 주로 진행되어 왔다(Ahlquist 등, 2003; Nagy 와 Pogany, 2011; Wang, 2015). 효모 시스템을 이용한 유전체 전반에 걸친 기주 인자 선발 연구를 통해 brome mosaic virus (BMV)와 tomato bushy stunt virus에서 각각 100개 이상의 기주 유전자가 바이러스의 감염에 영향을 끼친다는 것을 확인했다(Gancarz 등, 2011; Kushner 등, 2003; Nagy, 2016). BMV에서는 바이러스 게놈 번역 단계와 관련해서 기주 mRNA 번역에 관여하는 다양한 기주 단백질들(DED1, LSM1, LSM6, DHH1, PAT1)이 발견되었다(Diez 등, 2000; Kushner 등, 2003; Noueiry 등, 2000; Noueiry 등, 2003). 이 중에서 DED1 단백질은 효모에서 mRNA 번역 개시에 꼭 필요한 RNA helicase이며, 돌연변이가 일어났을 때 BMV의 RNA 중합효소(RNA-dependent RNA polymerase)의 번역을 방해하여 BMV의 증식을 막지만 효모의 세포 성장에는 영향을 주지 않는 것이 확인되었다(Hilliker 등, 2011; Noueiry 등, 2000).
일반적으로 바이러스는 효율적인 복제를 위해 VRC를 형성하고 세포 내 막 구조를 변형시켜 소구체(spherule)를 형성한다(de Castro 2013; den Boon 2010; Nagy 와 Pogany, 2011). 막의 기원은 바이러스에 따라 다르게 나타나며, VRC 형성과 막 구조 형성 과정에 바이러스 단백질 이외에 많은 기주 단백질들이 관여한다는 증거들이 많이 있다(Belov 와 van Kuppeveld, 2012; Nagy 와 Pogany, 2011). BMV에 의해 형성되는 막 구조도 효모 시스템에서 많은 연구가 진행되었으며, 소포체(endoplasmic reticulum, ER)에서 막 구조가 형성되는 것이 확인되었다(den Boon 2010; Diaz 와 Wang, 2014). 이러한 막 구성 요소와 유동성을 조절하거나 바이러스에 의한 막 구조 재배열의 유지에 관여하는 기주 단백질들은 BMV 증식에 필수적인 것으로 밝혀졌다(Diaz 와 Wang, 2014). Tobacco mosaic virus (TMV)의 경우에는 130K와 180K라는 복제 단백질들의 helicase 도메인과 ER의 막 관통 단백질인 TOM1이 상호작용하여 막 구조를 재배열하고 TMV의 증식을 촉진한다고 알려져 있다(Yamanaka 등, 2002).
바이러스의 세포 간 이동에도 바이러스 단백질과 상호작용하는 것으로 알려진 기주 유전자들이 중요하게 작용하고 있다. 바이러스는 식물체 전신 감염을 위해서 인접한 세포로 이동하기 위해 원형질연락사(plasmodesmata, PD)를 통과해야 한다 (Maule, 2008). 바이러스의 세포간 이동은 바이러스의 이동 단백질(movement protein, MP)과 다양한 기주 단백질들간의 상호작용으로 이루어지는데 바이러스 종류에 따라 외피 단백질(coat protein, CP)이 필요한 경우도 있다(Heinlein, 2015). TMV의 경우에는 TMV의 MP가 PD로 먼저 이동하여 바이러스 RNA-MP 복합체(vRNP)가 통과할 수 있도록 PD의 구멍 사이즈를 늘린다. TMV의 MP는 PD로 vRNP를 이동시키기 위해 미세소관을 이용하기 때문에 현재까지 알려진 대부분의 기주 단백질은 미세소관의 이동 및 형성과 관련이 있다(Heinlein, 2015). γ-튜불린과 미세소관의 조절 단백질인 EB1이 TBV의 MP와 직접적으로 상호작용하는 것으로 알려졌으며, 미오신 Ⅷ-1, -2, -B도 MP가 PD로 이동하는 데에 필수적인 것이 확인되었다(Amari 등, 2010, 2014; Ashby 등, 2006; Sambade 등, 2008). 실제로 미세소관의 역학과 관련된 단백질인 ATER2의 돌연변이체에서 TMV에 대한 감수성이 감소된 것을 확인했다(Ouko 등, 2010).
지금까지 밝혀진 바이러스 단백질과 상호작용하는 기주 인자들은 Yeast Two-hybrid 시스템 등 다양한 단백질 상호작용 연구를 통해 발견되었다. 그 중에서도 공동면역침강법(co-immunoprecipitation, Co-IP)을 통해 얻은 물질에 대한 질량분석(mass spectrometry)은 다양한 단백질간 상호작용을 밝히는 데 널리 쓰이는 방법 중에 하나이며, 방법적 간편성이 크고 적용성이 넓기 때문에 바이러스 단백질과 상호작용하는 기주 단백질을 탐색하는 데에 유용하게 쓰일 수 있다. 기본적으로 HA 또는 Flag와 같은 작은 항원결정인자(epitope)을 바이러스 단백질에 표지(tagging)하여 식물 세포에서 발현시킨 후, 해당 항원결정인자에 대한 항체를 이용하면 표지된 바이러스 단백질 및 이와 상호작용하는 기주 단백질을 Co-IP 할 수 있다. Co-IP를 통해 얻은 단백질 복합체를 liquid chromatography–tandem mass spectrometry (LC-MS/MS) 또는 matrix-assisted laser desorption ionization–time-of-flight mass spectrometer와 같은 질량분석 기법을 통해 분석하면 바이러스 단백질과 상호작용하는 기주 단백질을 선발할 수 있다(Ho 등, 2002; Mann 등, 2001; Vasilescu 와 Figeys, 2006). 표지된 바이러스 단백질을 식물 세포에서 발현하기 위해서는 아그로박테리움을 매개하는 T-DNA 전달 기술(agroinfiltration)이 주로 이용되고 있다(Annamalai 와 Rao, 2006; Lee 와 Yang, 2006; Vaghchhipawala 등, 2011). Agroinfiltration 기법은 T-DNA 구조 내에 35S 프로모터 등을 포함하여 손쉽게 원하는 유전자를 식물세포에서 과발현 시킬 수 있는 장점이 있다. 최근 한 연구에서는 바이러스의 감염성 DNA 클론을 이용하여 바이러스 유전자를 표지하고 식물 세포에서 표지된 단백질을 발현하여 Co-IP를 통해 기주 인자를 밝히기도 하였다(Seo 등, 2016). Soybean mosaic virus (SMV)의 감염성 DNA 클론을 이용하여 Flag으로 HC-Pro 유전자를 표지한 후 식물체를 감염시킨 다음, Co-IP와 LC-MS/MS를 통해 HC- Pro와 상호작용하는 기주 단백질을 선발할 수 있었다. HC-Pro는 바이러스 감염과정에서 다양한 역할을 하는 단백질로 알려졌으며, 단백질 가수분해 활성이 있어 자체 단백질의 C-말단을 자를 수 있다(Maia 등, 1996). 또한 SMV의 HC-Pro와 CP는 직접적으로 상호작용을 하여 바이러스 입자의 진딧물 전반에 관여하는 것으로 알려졌다(Blanc 등, 1997; Roudet 등, -Tavert 2002). 실제로 HC-Pro와의 Co-IP를 통해 얻어진 단백질로 CP 뿐만 아니라 기주인 콩의 GAPDH와 RetD가 새롭게 밝혀졌다(Seo 등, 2016). 이러한 바이러스 감염성 클론을 이용한 바이러스 유전자의 표지를 통한 Co-IP 기법은 실제 바이러스 감염 조건에서의 바이러스-기주 단백질간 상호작용을 밝힐 수 있다는 장점이 있으며, 손쉬운 클로닝으로 식물체 전신에서 표지된 유전자를 과발현 시킬 수 있다는 간편성이 있다.
새로운 바이러스 방제 전략을 세우거나 바이러스를 생물공학적으로 유용하게 이용하기 위해서 바이러스와 상호작용하는 기주 인자를 밝히는 연구는 지속적으로 중요하게 여겨지고 있다. 이렇게 밝혀진 기주 단백질 중에는 돌연변이가 일어나면 바이러스에 대한 열성 저항성을 유도하는 것들도 발견이 되었다(Lellis 등, 2002). 그러나 대부분의 기주 인자들은 식물의 생장, 유전자 발현, 신호전달에도 중요하게 작용하기 때문에, 이들에 인위적인 돌연변이를 일으킬 경우 정상적인 식물 생장을 저해할 수 있다(Clement 등, 2011; Jungkunz 등, 2011; Nicaise 등, 2007; Patrick 등, 2014; Ransom-Hodgkins, 2009; Zhang 등, 2015b). 따라서, 다양한 작용점에서의 기주 인자를 발굴하고 이들에 대한 정확한 작용기작을 이해하여 열성 저항성 획득을 위한 유전 소재로 개발할 필요가 있다.
유전자 교정 기술의 동향 및 열성 저항성 작물 개발에의 적용
바이러스와 상호작용하는 기주 인자를 찾으려는 많은 노력에도 불구하고 현재까지 밝혀진 기주 인자의 수는 많지 않다. 게다가, 특정 유전자를 찾은 후에도 작물에서 이를 대상으로 하여 기존의 무작위적 돌연변이 유발 방식과 교배육종으로 열성 저항성을 유도하기까지 상당한 시간이 걸린다. 그러나, 최근 CRISPR/Cas9 등 유전자 교정 기술의 급속한 발전으로 인해 손쉽게 특정 유전자에 대한 돌연변이 유발이 가능해짐에 따라, 이러한 기술을 이용한 병 저항성 식물 개발 연구에 있어 새로운 시대가 열리고 있다(Chandrasekaran 등, 2016; Pyott 등, 2016).
유전자 교정 기술은 site specific nuclease (SSN)를 이용하여 대상 유전자 또는 근처에서 이중 가닥의 절단(double strand break)을 기반으로 한다(Piatek 와 Mahfouz, 2017). SSN은 zinc finger nucleases (ZFNs), meganucleases, transcription activator-like effector nuclease (TALENs), clustered regularly interspaced palindromic repeats/CRISPR-associated 9 nuclease (CRISPR/Cas9)의 크게 4가지로 분류된다(Stella Montoya, 2016).
SSN에 의한 이중 가닥 절단은 두 가지 DNA 복구 체계를 통해 회복되는데, 하나는 non-homologous end-joining (NHEJ)이고 다른 하나는 homologous recombination이다(Ali 등, 2015; Aouida 등, 2014, 2015a, 2015b; Piatek 와 Mahfouz, 2017). NHEJ는 가장 간단하게 유전자의 knockout을 일으키는 기작으로(Barakate 와 Stephens, 2016), 종종 작은 insertion/deletion (INDEL)을 발생시키기도 한다(Wright 등, 2016). 이러한 INDEL 돌연변이는 대상 유전자의 코딩 또는 조절 서열에 영향을 미쳐 기능 상실(loss-of-function)의 결과를 나타나게 한다. Homologous recombination에 의한 이중 가닥 절단의 복구는 원하는 변형을 가진 DNA 복구 주형이 동시에 필요하기 때문에 NHEJ보다 더 복잡한 과정으로 발생한다(Stella 와 Montoya, 2016).
정교한 유전자 조작 기술은 ZFN을 이용하여 식물에서 처음으로 연구되었고, 이어서 meganucleases, TALEN을 이용한 연구들이 수행되었다(Cheng 등, 2015). ZEN과 TALEN은 베고모바이러스에 대한 저항성 작물 개발에 이용된 바 있지만, 대상 서열에 결합하는 DNA 결합 단백질 제작에 많은 비용과 시간이 소요되는 단점이 있다. 이에 반해, CRISPR/Cas9 기술은 특정 서열과 결합하는 단백질을 일일이 제작해야하는 ZEN과 TALEN 기술과 다르게 guide RNA를 통해 목표 유전자를 특정할 수 있기 때문에 훨씬 쉽게 다룰 수 있다는 장점이 있다(Doudna 와 Charpentier, 2014; Quetier, 2016).
CRISPR/Cas9는 외부 핵산의 침입을 막기 위해 원핵 생물이 사용하는 면역 시스템을 이용한 방법이다(Barrangou 와 Marraffini, 2014). 박테리아나 고세균은 획득한 작은 조각의 외래 핵산이나 spacer를 자신의 유전체에 삽입하고 있다가 재차 감염 시 이 작은 조각들을 발현하여 외래 유전자를 특정하고 Cas9 endonuclease가 정확히 작동할 수 있도록 돕는다. CRISPR/Cas9은 protospacer-associated motif 근처의 목표 서열을 매우 정확하게 자르는데, 현재 단순한 미생물부터 복잡한 식물이나 동물에 이르기까지 많은 종에서 성공적으로 사용되고 있다(Zetsche 등, 2015).
다양한 방식의 유전자 교정 기술은 모델 식물뿐만 아니라 실제 여러 작물에도 적용되고 있다. 특히 작물 육종 측면에서 유전자 교정 기술의 가장 뛰어난 장점은 유전자 교정 후 다음 세대의 유전자형 분리(segregation)를 통해 유전자 교정을 위해 삽입된 transgene을 제거할 수 있다는 점이다(Fig. 2). 최근에 이러한 CRISPR/Cas9 시스템을 이용하여 기주 유전자인 eIFi-so4E에 대한 돌연변이를 유발하여 turnip mosaic virus에 대한 열성 저항성 돌연변이 식물을 제작했다는 보고가 있다(Pyott 등, 2016). 또한, 오이(Cucumis sativus)에서도 CRISPR/Cas9 시스템을 이용하여 eIF4E 유전자에 돌연변이를 유발시켜 cucumber vein yellowing virus와 두 종의 포티바이러스에 대한 열성 저항성이 유도된 식물이 제작된 바 있다 (Chandrasekaran 등, 2016). 다배체인 콩에서는 ZFN을 사용하여 병저항성 돌연변이체를 만든 기록이 있으며, 이질 육배체인 밀에서도 TALEN을 이용하여 3가지의 MILDEW RESISTANCE LOCUS 유전자에 동시에 돌연변이를 유발시켜 흰가루병을 일으키는 곰팡이에 대한 열성 저항성을 유도했다(Curtin 등, 2011; Wang 등, 2014). 최근까지의 이러한 CRISPR/Cas9 기반의 기술은 대상 유전자 또는 그 유전자의 프로모터에 INDEL을 유발하여 knockout시키는 것으로 열성 저항성을 유도하였다. 이러한 knockout 방법은 과정이 상대적으로 간단하며, 원하는 돌연변이를 만들기 쉽고, 작물 육종에 편리하다는 장점이 있지만 대상 유전자가 식물 생장 및 발달에 필수적이라면 적용이 불가능할 수 있다. 매우 최근에는 더욱 향상된 CRISPR/Cas9 기술로서 특정 아미노산에 돌연변이를 유입하는 단일 염기 교정(base editing)이 가능해지고 있다(Cermak 등, 2017; Zong 등, 2017). 이러한 단일 염기 교정 기술을 이용하면 바이러스 단백질과의 상호작용에 중요한 기주 단백질의 아미노산에 돌연변이를 유입함으로써, 바이러스 단백질과의 상호작용을 억제하여 열성 저항성을 유도할 수 있을 것이다. 이러한 접근법은 특정 유전자의 knockout으로 발생할 수 있는 식물 생장에의 악영향을 최소화할 수 있으며, 자연적으로 발생하는 열성 저항성 돌연변이와 유사하기 때문에 안전하고 친환경적이라 할 수 있다. 아직까지는 CRISPR/Cas9 기술로 유도된 열성 저항성이 계속 유지되는지 지켜봐야 할 필요가 있으며, 부정확한 변이(off-target) 발생의 위험성이 존재하고 있으므로 기술의 정교성 향상을 위한 지속적인 연구가 필요한 상황이다. 또한 식물의 형질 개선을 위해서는 유전자 교정 기술의 발전과 더불어 이러한 기술을 적용할 유전 소재 발굴이 매우 중요한 시점이다. 앞으로 다각도의 연구를 통해 다양한 열성 저항성 유전 소재의 발굴이 이루어져야 하며, 향상된 유전자 교정 기술을 바탕으로 안정적으로 열성 저항성이 유도된 작물을 개발하려는 노력이 필요할 것이다.
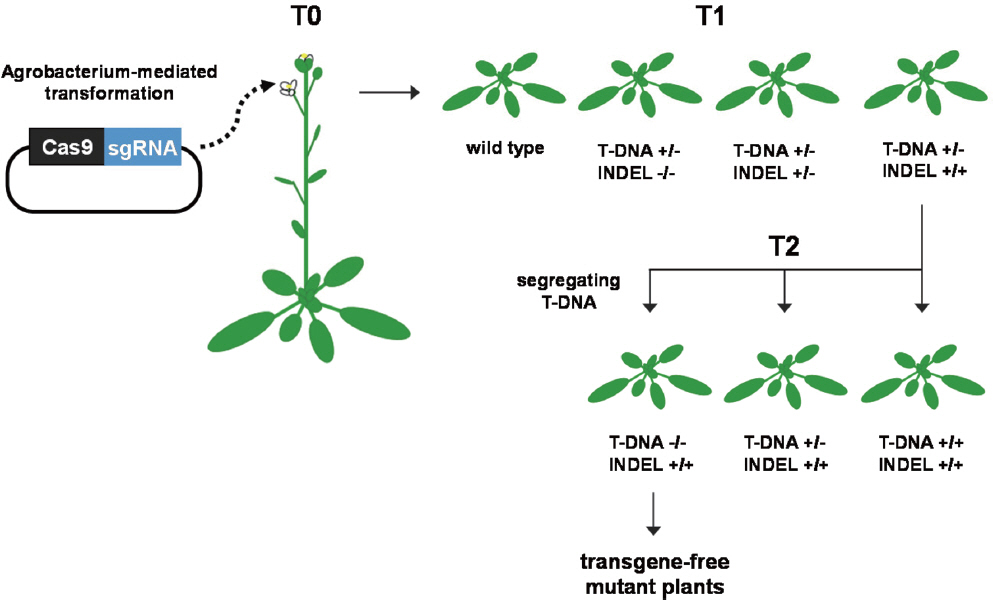
Simplified overview for generation of transgene-free, genome-edited plants using CRISPR/Cas9. Arabidopsis plants can be transformed by Agrobacterium carrying a CRISPR/Cas9 construct. Targeted mutagenesis events can occur in transgenic T1 plants. In T2 plants, the T-DNA segregates in a Mendelian fashion for single-locus lines, and thereby transgene-free, homozygously mutated plants can occur in T2.
결론 및 제언
식물에서의 바이러스병에 대한 열성 저항성은 바이러스와 기주 식물 간의 분자적 상호작용을 기반으로 이루어진다. 바이러스의 병원성인자와 상호작용하는 기주 유전자를 발굴하면 저항성 품종 육성을 위한 유전 소재로의 활용이 가능해질 수 있다. 최근 CRISPR/Cas9 등 유전자 교정 기술의 급속한 발전으로 바이러스 단백질과 상호작용하는 기주 유전자에 대한 정교한 유전자 교정을 통해 바이러스 열성 저항성 작물을 개발할 수 있게 되었고, 조금씩 이 기술을 적용한 작물 개발 연구가 시작되고 있다. 바이러스와 열성 저항성 유전자에 대한 분자유전학적인 기초 연구와 함께 유전자 교정 기술과 같은 새로운 생명공학 기술을 이용한 응용 연구의 결합은 바이러스병 방제와 같이 기존 기술로 해결이 어려운 농업적 요구에 효과적으로 대응할 수 있는 새로운 방안을 제시할 수 있을 것으로 보인다.
요 약
식물 바이러스는 작물 생산량 손실을 일으키는 주요 병원체 중 하나로, 돌연변이 발생이 빈번하고 치료 약제가 개발되어 있지 않아 방제가 매우 어렵다. 이러한 바이러스병을 방제하기 위한 가장 효과적인 방법은 저항성 품종을 재배하는 것이며, 바이러스 저항성 품종을 개발하기 위해서는 바이러스와 기주 식물 간의 다양한 유전자적 상호작용에 대한 정확한 이해가 필요하다. 열성 저항성은 병원체가 살아가는 데에 필요한 식물 유전자가 결핍되었을 때 획득되는데, 저항성 유전자(R gene)에 의해 유도되는 우성 저항성에 비해 넓은 범위의 저항성을 발현하고 돌연변이 출현에 쉽게 저항성이 깨지지 않는 특성을 보인다. 현재까지 알려진 바이러스병에 대한 열성 저항성 유전자는 대부분 순행유전학(forward genetics)를 통해 밝혀졌으나, 최근 CRISPR/Cas9 등을 이용한 유전자 교정 기술의 급속한 발전에 힘입어 역유전학(reverse genetics)을 통한 열성 저항성 작물 개발의 가능성이 열리고 있다. 이러한 역유전학적 접근을 통한 열성 저항성 작물 개발은 먼저 바이러스 단백질과 상호작용하는 기주 인자를 밝히고 이들간의 상호작용을 억제하도록 하는 기주 인자에 대한 유전자 교정을 통해 이루어 질 수 있다. 본 논문에서는 열성 저항성에 대한 소개와 새로운 열성 저항성 후보 유전 소재 발굴을 위한 기주 인자 연구의 중요성 및 방법을 소개하고, 열성 저항성 작물 개발에 적용할 수 있는 유전자 교정 기술의 최신 동향에 관해 정리하였다.
Conflicts of Interest
No potential conflict of interest relevant to this article was reported.
Acknowledgements
This research was supported in part by grants from the Next-Generation BioGreen 21 Program (PJ013129) funded by the Rural Development Administration of Korea and the Young Researcher Program (2018R1C1B5029927) funded by the National Research Foundation of Korea.