

ņä£ ļĪĀ
ļöĖĻĖ░ļłäļźĖņśżĻ░łļ░öņØ┤ļ¤¼ņŖż(Strawberry mild yellow edge virus, SMYEV)ļŖö 1922ļģäņŚÉ ļ»ĖĻĄŁ ņ║śļ”¼ĒżļŗłņĢäņŚÉņä£ ņ▓śņØīņ£╝ļĪ£ ļ│┤Ļ│ĀļÉ£ ņØ┤Ēøä Ēśäņ×¼ ņĀäņäĖĻ│äņŚÉņä£ Ļ░Ćņן ļäÉļ”¼ ļ░£ļ│æĒĢśĻ│Ā ņ׳ļŖö ļöĖĻĖ░ ļ░öņØ┤ļ¤¼ņŖż ņżæ ĒĢśļéśņØ┤ļŗż(Horne, 1922; Jelkmann ļō▒, 1990; MartinĻ│╝ Tzanetakis, 2006; ThompsonĻ│╝ Jelkmann, 2004; TzanetakisņÖĆ Martin, 2013). SMYEVļŖö ļČäļźśĒĢÖņĀüņ£╝ļĪ£ AlphaflexiviridaeĻ│╝ņØś Potexvirus ņåŹņŚÉ ņåŹĒĢśļ®░, ņ£ĀņØ╝ĒĢśĻ▓ī ņ¦äļö¦ļ¼╝ņŚÉ ņØśĒĢ┤ ņĀäņŚ╝ĒĢśļŖö PotexvirusļĪ£ ļöĖĻĖ░ņ¦äļö¦ļ¼╝ņØĖ Chaetosiphon fragaefoliiņŚÉ ņØśĒĢ┤ ņśüņåŹ ņĀäņŚ╝ļÉ£ļŗż(Jelkmann ļō▒, 1990). SMYEVņØś Ļ░ÉņŚ╝ņ”ØņāüņØĆ ļöĖĻĖ░ņØś ĒÆłņóģ ļ░Å ļ░öņØ┤ļ¤¼ņŖż ļ│ĆņØ┤ņŻ╝(strain)ņŚÉ ļö░ļØ╝ ļ│æņ¦Ģ ļ░Å Ēö╝ĒĢ┤ ņĀĢļÅäĻ░Ć ļŗżļź┤ļŗżĻ│Ā ņĢīļĀżņĀĖ ņ׳ļŗż. MartinĻ│╝ Tzanetakis (2006)ņØś ļ│┤Ļ│ĀņŚÉ ļö░ļź┤ļ®┤ Ļ░Éņłśņä▒ ĒÆłņóģņØś Ļ▓ĮņÜ░ ņ×Ä ļÆżĒŗĆļ”╝ņØ┤ļéś ņ£äņČĢņ”Øņāü ļ░Å ņ×Ä Ļ░Ćņןņ×Éļ”¼Ļ░Ć ĒÖ®ĒÖöļÉśļŖö ņ”ØņāüņØä ļ│┤ņØ┤ņ¦Ćļ¦ī ļé┤ņä▒ ĒÆłņóģņØś Ļ▓ĮņÜ░ ņĀäĒśĆ ļ│æņ¦ĢņØ┤ ļéśĒāĆļéśņ¦Ć ņĢŖĻĖ░ļÅä ĒĢ┤ ņłśĒÖĢļ¤ē ļ░Å ņāØņ£Ī ņØ┤ņāüĻ│╝ Ļ░ÖņØĆ Ēö╝ĒĢ┤ņ£©ļÅä 0-30%ļĪ£ ļ¦żņÜ░ ņāüņØ┤ĒĢśĻ▓ī ļéśĒāĆļé£ļŗż. ĻĄŁļé┤ņØś Ļ▓ĮņÜ░ 2009ļģäĻ│╝ 2011ļģäņŚÉ ļöĖĻĖ░ņŚÉņä£ SMYEVĻ░Ć ļ│┤Ļ│ĀļÉ£ ļ░ö ņ׳ļŖöļŹ░, ļöĖĻĖ░ ĒÆłņóģņŚÉ ļö░ļØ╝ ņ×Ä Ļ░Ćņןņ×Éļ”¼ņŚÉ ņŻ╝ļ”ä ļśÉļŖö ĒÖ®ĒÖö, ņ×ÄņŚÉ ļ¬©ņ×ÉņØ┤Ēü¼ ņ”ØņāüņØ┤ ļéśĒāĆļéśņ¦Ćļ¦ī ļ¼┤ņ”ØņāüņŚÉņä£ļÅä ļ░öņØ┤ļ¤¼ņŖżĻ░Ć Ļ▓ĆņČ£ļÉśņ¢┤ ļ░öņØ┤ļ¤¼ņŖż Ļ░ÉņŚ╝Ļ│╝ ļ│æņ¦ĢņØś ņŚ░Ļ┤Ćņä▒ņØĆ ņĀüņØĆ Ļ▓āņ£╝ļĪ£ ļ│┤Ļ│ĀĒĢśņśĆļŗż(Cho ļō▒, 2011; Choi ļō▒, 2009).
SMYEVļŖö ņĄ£ņ┤ł ļ░£ņāØ ļŗ╣ņŗ£ņŚÉļŖö ĻĄ¼ĒśĢļ░öņØ┤ļ¤¼ņŖżņØĖ LuteovirusļĪ£ ļČäļźśļÉśņŚłņ£╝ļéś(Plakidas, 1927), ņØ┤Ēøä MY18Ļ│╝ D74 ļČäļ”¼ņŻ╝(isolate)ņØś ņĀäņ▓┤Ļ▓īļåłņØ┤ Ļ▓░ņĀĢļÉśĻ│Ā ņ£ĀņĀäņ×É ļ╣äĻĄÉ ļČäņäØņØä ĒåĄĒĢ┤ PotexvirusļĪ£ ļČäļźśļÉśņŚłĻ│Ā ļ░öņØ┤ļ¤¼ņŖż ņ×ģņ×ÉņØś ĒśĢĒā£ļÅä 480├Ś13 nmņØś ņé¼ņāüĒśĢ ņ×ģņ×Éņ×äņØ┤ ĒÖĢņØĖļÉśņŚłļŗż(Jelkmann ļō▒, 1990). Ēśäņ×¼Ļ╣īņ¦Ć SMYEV ņĀäņ▓┤ Ļ▓īļåł ņŚ╝ĻĖ░ņä£ņŚ┤ņØ┤ ļ░ØĒśĆņ¦ä 8Ļ░£ ļČäļ”¼ņŻ╝(MY18, D74, Berra 2, AB41-01, AB41-02, NB1165, NS26, AB5-2)ņŚÉ ļö░ļź┤ļ®┤ SMYEVļŖö 5-6Ļ░£ņØś ORFļź╝ ļ░£ĒśäĒĢśļ®░ Ēü¼ĻĖ░ļŖö 5.9-7 kbņØś ļŗ©Ļ░Ćļŗź RNA Ļ▓īļåł ļ░öņØ┤ļ¤¼ņŖżņ×äņØ┤ ĻĄ¼ļ¬ģļÉśņŚłļŗż(Bhagwat ļō▒, 2015; LamprechtĻ│╝ Jelkmann, 1997; Ma ļō▒, 2015; ThompsonĻ│╝ Jelkmann, 2004). ņ£ĀņĀäņ×É ļČäņäØņØĆ Ēśäņ×¼ ļ░öņØ┤ļ¤¼ņŖżņØś ņóģ ļČäļźśņŚÉ Ļ░Ćņן ļäÉļ”¼ ņØ┤ņÜ®ļÉśļŖö ļ░®ļ▓Ģņ£╝ļĪ£ PotexvirusņØś Ļ▓ĮņÜ░ ņŻ╝ļĪ£ RNA ņżæĒĢ®ĒÜ©ņåī(RNA-dependent RNA polymerase)ņÖĆ ņÖĖĒö╝ļŗ©ļ░▒ņ¦ł(Coat protein, CP) ņ£ĀņĀäņ×ÉņØś ļ╣äĻĄÉļČäņäØņØä ĒåĄĒĢ┤ strainņØ┤ļéś variantsļź╝ ĻĄ¼ļČäĒĢ£ļŗż(King ļō▒, 2012). ņ£ĀņĀäņ×É ļČäņäØņŚÉ ĻĘ╝Ļ▒░ĒĢ£ Ļ│äĒåĄĒĢÖņĀü ļČäņäØņØĆ ļ░öņØ┤ļ¤¼ņŖżņØś ņ¦äĒÖö ļ░Å ņ£ĀņĀäņĀü ļ│ĆņØ┤ņ¢æņāüņØä ĒīīņĢģĒĢśļŖöļŹ░ ļ¦żņÜ░ ņżæņÜöĒĢ£ ņĀĢļ│┤Ļ░Ć ļÉśĻ│Ā ņ׳ļŗż. SMYEVļÅä Ļ│äĒåĄĒĢÖņĀü ļČäņäØņØä ĒåĄĒĢ┤ ņ£ĀņĀäņĀüņ£╝ļĪ£ ĒÖĢņŚ░Ē׳ ļŗżļźĖ 3Ļ░£ņØś ļ│ĆņØ┤ņ¦æļŗ©ņØ┤ ņĪ┤ņ×¼ĒĢśļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ņŚłņ£╝ļéś(ThompsonĻ│╝ Jelkmann, 2004), ņĄ£ĻĘ╝ ņĢäļź┤ĒŚ©Ēŗ░ļéśņÖĆ ņ║ÉļéśļŗżņŚÉņä£ ļČäļ”¼ļÉ£ SMYEV ļČäļ”¼ņŻ╝ņŚÉ ļīĆĒĢ£ ļČäņäØ Ļ▓░Ļ│╝ 2Ļ░£ņØś ņāłļĪ£ņÜ┤ ļ│ĆņØ┤ņ¦æļŗ©ņØ┤ ļŹö ņĪ┤ņ×¼ĒĢśļŖö Ļ▓āņØ┤ ļ░ØĒśĆņĀĖ ļ░öņØ┤ļ¤¼ņŖż ļ│ĆņØ┤ņŻ╝Ļ░ä ļŗżņ¢æņä▒ņØ┤ ļŹö ņ¦äĒÖöļÉśĻ│Ā ņ׳ļŖö Ļ▓āņ£╝ļĪ£ ņČöņĖĪļÉ£ļŗż (Bhagwat ļō▒, 2015; Ma ļō▒, 2015; Torrico ļō▒, 2016).
ļ│Ė ļģ╝ļ¼ĖņŚÉņä£ļŖö Cho ļō▒(2011)ņŚÉ ņØśĒĢ┤ ĻĖ░ņĪ┤ņŚÉ ļ│┤Ļ│ĀļÉśņŚłļŹś KNS1 ļČäļ”¼ņŻ╝ņÖĆ ņĄ£ĻĘ╝ ĻĄŁļé┤ ņ£Īņä▒ ļöĖĻĖ░ ĒÆłņóģņŚÉņä£ ļČäļ”¼ļÉ£ 2Ļ░£ņØś SMYEV ĻĄŁļé┤ ļČäļ”¼ņŻ╝ļź╝ ļŹö ĒÖĢļ│┤ĒĢśņŚ¼ ņØ┤ļōżņØś CP ņ£ĀņĀäņ×É ņŚ╝ĻĖ░ņä£ņŚ┤ņØä ļČäņäØĒĢśĻ│Ā, ĻĖ░ņĪ┤ NCBIņŚÉ ļō▒ļĪØļÉ£ SMYEVņØś 48Ļ░£ ļČäļ”¼ņŻ╝ļōżĻ│╝ņØś Ļ│äĒåĄĒĢÖņĀü ļČäņäØņØä ĒåĄĒĢ┤ ĻĄŁļé┤ SMYEV ļČäļ”¼ņŻ╝ļōżņØś ņ£ĀņĀäņĀü ļŗżņ¢æņä▒ņŚÉ ļīĆĒĢ£ Ļ▓░Ļ│╝ļź╝ ļ│┤Ļ│ĀĒĢśĻ│Āņ×É ĒĢ£ļŗż.
ņ×¼ļŻī ļ░Å ļ░®ļ▓Ģ
ļ░öņØ┤ļ¤¼ņŖż ļČäļ”¼ņŻ╝ ĒÖĢļ│┤ ļ░Å ņÖĖĒö╝ļŗ©ļ░▒ņ¦ł ļČäņäØ
ĻĄŁļé┤ ņ£Īņä▒ ļöĖĻĖ░ ĒÆłņóģ(ļ¦żĒ¢ź, ņäżĒ¢ź, ĻĖłņŗż, ņŗ╝ĒāĆ, Ļ│ĀĒĢś, Ļ░ÉĒÖŹ, ļ»ĖĒÖŹ, ņןĒĢś, ļ¼┤ĒĢś)ņØä ļīĆņāüņ£╝ļĪ£ SMYEV Ļ░ÉņŚ╝ ņŚ¼ļČĆļź╝ ņĪ░ņé¼ĒĢśņśĆļŗż. 2017ļģä 2ņøöļČĆĒä░ 2018ļģä 8ņøöĻ╣īņ¦Ć ļ¦żĒ¢ź, ĻĖłņŗż, ņäżĒ¢ź, ņŗ╝ĒāĆ ĒÆłņóģņØĆ Ļ▓Įļé© ņ¦äņŻ╝ņÖĆ ņČ®ļé© ļģ╝ņé░ ļ░Å ļČĆņŚ¼ņŚÉņä£ ņĪ░ņé¼ĒĢśņśĆĻ│Ā, Ļ│ĀĒĢś, Ļ░ÉĒÖŹ, ļ»ĖĒÖŹ, ņןĒĢś, ļ¼┤ĒĢśĒÆłņóģņØĆ Ļ░ĢņøÉļÅä ĒÅēņ░Į ļ░Å Ļ│ĀļĀ╣ņ¦ĆļåŹņŚģņŚ░ĻĄ¼ņåī ļé┤ ĒżņןņŚÉņä£ ņĪ░ņé¼ĒĢśņśĆļŗż. SMYEV ņ¦äļŗ©ņØĆ CP ņ£ĀņĀäņ×Éļź╝ ĒżĒĢ©ĒĢśļŖö ĒŖ╣ņØ┤ ĒöäļØ╝ņØ┤ļ©Ė (SMYEV-FOR: 5ŌĆÖ-ACAATCGCCCTGGTCAGTAATTCC-3ŌĆÖ, SMYEVREV: 5ŌĆÖ-TGGAGCTGGGTAGAAGAAGTGAGA-3ŌĆÖ)ļź╝ ņØ┤ņÜ®ĒĢśņśĆļŗż. ņ£ĀņĀäņ×É ņ¦äļŗ©ņØä ņ£äĒĢ£ RNA ņČöņČ£ņØĆ TransZol (TransGen Biotech, China)ņØä ņØ┤ņÜ®ĒĢśņśĆĻ│Ā, SuPrimeScript RT-Premix (GeNet Bio, Korea)ļĪ£ 50┬░C/30 m; 98┬░C/30 s; <98┬░C/10 s, 60┬░C/30 s, 68┬░C/1 m> 35 cycles; 68┬░C/5 mņØś ņĪ░Ļ▒┤ņ£╝ļĪ£ RT-PCRĒĢśņśĆļŗż. PCR ņé░ļ¼╝ņØĆ ņĀäĻĖ░ņśüļÅÖņŚÉņä£ ĒÖĢņØĖ Ēøä, QIAquick PCR purification kit (Qiagen, Germany)ļĪ£ ņĀĢņĀ£ĒĢśņŚ¼ pGEM-T easy vector(Promega, USA)ņŚÉ Ēü┤ļĪ£ļŗØĒĢśĻ│Ā ņÖĖĒö╝ļŗ©ļ░▒ņ¦ł ņŚ╝ĻĖ░ņä£ņŚ┤ņØä Ļ▓░ņĀĢĒĢśņśĆļŗż. ĻĘĖ Ļ▓░Ļ│╝, ņäżĒ¢ź(ņĢĮĒĢ£ ļ¬©ĒŗĆ)Ļ│╝ Ļ░ÉĒÖŹ(ņ×Ä Ļ░Ćņןņ×Éļ”¼ ļ│Ćņāē) ĒÆłņóģņŚÉņä£ SMYEVņØś Ļ░ÉņŚ╝ņØä ĒÖĢņØĖĒĢśĻ│Ā ņĀäņ×ÉĒśäļ»ĖĻ▓Įņ£╝ļĪ£ ņé¼ņāüĒśĢ ņ×ģņ×Éļź╝ ĒÖĢņØĖĒĢśņśĆļŗż. Ļ░üĻ░üņØś ļČäļ”¼ņŻ╝ļŖö SMYEV-SHņÖĆ SMYEV-KHņ£╝ļĪ£ ļ¬ģļ¬ģĒĢśĻ│Ā ņĀäņ▓┤ CP ņ£ĀņĀäņ×Éļź╝ NCBIņŚÉ ļō▒ļĪØĒĢśņśĆļŗż.
Ļ│äĒåĄĒĢÖņĀü ņ£ĀņŚ░Ļ┤ĆĻ│ä ļ░Å ņ£ĀņĀäņĀü ļŗżņ¢æņä▒ ļČäņäØ
ĻĄŁļé┤ņŚÉņä£ ļČäļ”¼ļÉ£ SMYEVņØś CPņŚÉ ļīĆĒĢ£ Ļ│äĒåĄĒĢÖņĀü ņ£ĀņĀäņ×É ļČäņäØņØä ņ£äĒĢ┤ NCBIņŚÉ ļō▒ļĪØļÉ£ SMYEV CP ņŚ╝ĻĖ░ņä£ņŚ┤ ņĀĢļ│┤ļź╝ ņłśņ¦æĒĢśņśĆļŗż(Table 1). ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ļČäļ”¼ļÉ£ SHņÖĆ KH, ĻĖ░ņĪ┤ņŚÉ ļČäļ”¼ļÉ£ KNS1 ĻĄŁļé┤ ļČäļ”¼ņŻ╝ļź╝ ĒżĒĢ©ĒĢśņŚ¼ ņ┤Ø 48Ļ░£ņØś ļČäļ”¼ņŻ╝ņŚÉ ļīĆĒĢ£ Ļ│äĒåĄĒĢÖņĀü ņ£ĀņŚ░Ļ┤ĆĻ│äļź╝ ļČäņäØĒĢśņśĆļŗż. ņŚ╝ĻĖ░ņä£ņŚ┤ ņĀĢļĀ¼ņØĆ Clustal X2 ĒöäļĪ£ĻĘĖļשņØä ņØ┤ņÜ®ĒĢśņśĆĻ│Ā Phylogenetic treeļŖö Mega 6ņØś Neighbor-joining (NJ) ļ░®ņŗØņ£╝ļĪ£ ņ×æņä▒ĒĢśņśĆļŗż. NJ-treeņØś ĒåĄĻ│äņĀü ņ£ĀņØśņä▒ņØĆ 1000ĒÜī ļ░śļ│ĄņŗżĒ¢ēņØś bootstrapņØä ņĀüņÜ®ĒĢśņŚ¼ ļČäņäØĒĢśņśĆļŗż. Potato virus XņØś CPņ£ĀņĀäņ×É(NC011620)ļŖö outgroupņ£╝ļĪ£ ņØ┤ņÜ®ĒĢśņśĆļŗż. SMYEVņØś ņ£ĀņĀäņ×É ļŗżņ¢æņä▒ ļČäņäØņØä ņ£äĒĢ┤ DnaSP (version 5)ņØä ņØ┤ņÜ®ĒĢśņŚ¼ phylogenetic treeņŚÉ ņØśĒĢ┤ Ļ│äĒåĄņØ┤ ļČäļźśļÉ£ Ļ░ü subgroup ļé┤ ļ░Å subgroup Ļ░ä ņ£ĀņĀäĻ▒░ļ”¼ ļ░Å ņŚ╝ĻĖ░ņä£ņŚ┤ ļŗżņ¢æņä▒ņØś ņÜöņØĖļōż(number of monomorphic sites, number of polymorphic site, number of mutations, singleton variable sites, parsimony informative site, synonymous substitutions, nonsynonymous substitutions)ņØä Ļ▓░ņĀĢĒĢśņśĆļŗż(LibradoņÖĆ Rozas, 2009).
Table┬Ā1
Information of SMYEV isolates available in the NCBI compared in this study
| ŌĆāŌĆāIsolate | ŌĆāŌĆāHost | Year | ŌĆāCountry | ŌĆāAcc. No. | ŌĆāŌĆāReference |
|---|---|---|---|---|---|
| SH | F. x ananassa cv. Seolhyang | 2017 | Korea | MG418838 | In this study |
| KH | F. x ananassa cv. Kamhong | 2018 | Korea | MK040457 | In this study |
| KNS1 | F. x ananassa cv. Seolhyang | 2011 | Korea | EU284709 | Cho et al., 2011 |
| 1CH | F. chiloensis | 2001 | Chile | AJ577337 | Thompson and Jelkmann, 2004 |
| 2CH | F. chiloensis | 2001 | Chile | AJ577338 | Thompson and Jelkmann, 2004 |
| 3CH | F. chiloensis | 2001 | Chile | AJ577339 | Thompson and Jelkmann, 2004 |
| 4CH | F. chiloensis | 2001 | Chile | AJ577340 | Thompson and Jelkmann, 2004 |
| 5CH | F. chiloensis | 2001 | Chile | AJ577341 | Thompson and Jelkmann, 2004 |
| 6CH | F. chiloensis | 2001 | Chile | AJ577342 | Thompson and Jelkmann, 2004 |
| 7CH | F. chiloensis | 2001 | Chile | AJ577343 | Thompson and Jelkmann, 2004 |
| 10CH | F. chiloensis | 2001 | Chile | AJ577346 | Thompson and Jelkmann, 2004 |
| D/L.9 | F. x ananassa cv. Elsanta | 2002 | Germany | AJ577344 | Thompson and Jelkmann, 2004 |
| D/L.13 | F. x ananassa cv. Elsanta | 2002 | Germany | AJ577347 | Thompson and Jelkmann, 2004 |
| D/L.14 | F. x ananassa cv. Elsanta | 2002 | Germany | AJ577348 | Thompson and Jelkmann, 2004 |
| D/L.19 | F. x ananassa cv. Elsanta | 2002 | Germany | AJ577349 | Thompson and Jelkmann, 2004 |
| D/M.110 | F. x ananassa cv. Elsanta | 2003 | Germany | AJ577352 | Thompson and Jelkmann, 2004 |
| D/K.159 | F. x ananassa cv. Elsanta | 2003 | Germany | AJ577353 | Thompson and Jelkmann, 2004 |
| D/V.180 | F. x ananassa cv. Elsanta | 2003 | Germany | AJ577354 | Thompson and Jelkmann, 2004 |
| D74 | F. vesca clone UC5 | 1974 | Germany | AJ577359 | Thompson and Jelkmann, 2004 |
| MY18 | F. x ananassa cv. Totem | 1970s | USA | D12517 | Thompson and Jelkmann, 2004 |
| 1182-53F | F. x ananassa selection 1187 | 1994 | USA | AJ577355 | Thompson and Jelkmann, 2004 |
| WSU1988 | F. x ananassa selection WSU1988 | 1988 | USA | AJ577358 | Thompson and Jelkmann, 2004 |
| 9Redland | F. x ananassa cv. Redlands crimson | 1996 | Australia | AJ577345 | Thompson and Jelkmann, 2004 |
| 69N | F. x ananassa cv. unknown | 1996 | Belgium | AJ577350 | Thompson and Jelkmann, 2004 |
| IndukaA | F. vesca clone UC5 | 2001 | Czech Republic | AJ577356 | Thompson and Jelkmann, 2004 |
| IndukaB | F. vesca clone UC5 | 2001 | Czech Republic | AJ577357 | Thompson and Jelkmann, 2004 |
| 314CP2cav | F. x ananassa cv. Elsanta | 2002 | Italy | AJ577351 | Thompson and Jelkmann, 2004 |
| sy01 | NA | NA | China | AY955375 | unpublished |
| sy02 | NA | NA | China | EU107084 | unpublished |
| sy03 | NA | NA | China | EU107085 | unpublished |
| sy04 | NA | NA | China | EU107086 | unpublished |
| 13-2-5 | F. x ananassa cv. Camarosa | 2008 | Argentina | KP284152 | Torrico et al., 2016 |
| 13-3-4 | F. x ananassa cv. Camarosa | 2008 | Argentina | KP284153 | Torrico et al., 2016 |
| 15 | F. x ananassa cv. Camarosa | 2007 | Argentina | KP284154 | Torrico et al., 2016 |
| 16-1 | F. x ananassa cv. Camarosa | 2007 | Argentina | KP284162 | Torrico et al., 2016 |
| 16-2 | F. x ananassa cv. Camarosa | 2007 | Argentina | KP284155 | Torrico et al., 2016 |
| 16-4-5 | F. x ananassa cv. Camarosa | 2007 | Argentina | KP284156 | Torrico et al., 2016 |
| 36-1-3 | F. x ananassa cv. Albion | 2010 | Argentina | KP284157 | Torrico et al., 2016 |
| 36-2-4-6 | F. x ananassa cv. Albion | 2010 | Argentina | KP284158 | Torrico et al., 2016 |
| 53 | F. x ananassa cv. Camarosa | 2008 | Argentina | KP284159 | Torrico et al., 2016 |
| 264 | F. x ananassa cv. Albion | 2010 | Argentina | KP284160 | Torrico et al., 2016 |
| 20 | F. x ananassa cv. Camarosa | 2005 | Argentina | KP284161 | Torrico et al., 2016 |
| Berra2 | F. x ananassa cv. Camarosa | 2011 | Argentina | KX150372 | Torrico et al., 2016 |
| AB41-01 | F. x ananassa | 2013 | Canada | KR350470 | Bhagwat et al., 2015 |
| AB41-02 | F. x ananassa | 2013 | Canada | KR350471 | Bhagwat et al., 2015 |
| NB1165 | F. x ananassa | 2014 | Canada | KR559735 | Bhagwat et al., 2015 |
| NS26 | F. x ananassa | 2014 | Canada | KR559736 | Bhagwat et al., 2015 |
| AB5-2 | F. x ananassa | 2014 | Canada | KR707814 | Ma et al., 2015 |
Ļ▓░Ļ│╝ ļ░Å Ļ│Āņ░░
SMYEV ņÖĖĒö╝ļŗ©ļ░▒ņ¦łņØś ĻĄŁļé┤ ļČäļ”¼ņŻ╝ ņŚ╝ĻĖ░ņä£ņŚ┤ ļČäņäØ
ĻĄŁļé┤ņŚÉņä£ ņāłļĪ£ ļČäļ”¼ļÉ£ SMYEV 2Ļ░£ ļČäļ”¼ņŻ╝ SHņÖĆ KHņØś CP ņ£ĀņĀäņ×ÉņØś ņĀäņ▓┤ ņŚ╝ĻĖ░ņä£ņŚ┤ņØä Ļ▓░ņĀĢĒĢśņŚ¼ Ļ░üĻ░ü MG418838, MK040457ļĪ£ GenBankņŚÉ ļō▒ļĪØĒĢśņśĆļŗż. SHņÖĆ KH ļČäļ”¼ņŻ╝ņØś CP ņĀäņ▓┤ ņŚ╝ĻĖ░ņä£ņŚ┤ņØś ĻĖĖņØ┤ļŖö 729 bpļĪ£ ņóģĻ▓░ņĮöļÅłņØä ņĀ£ņÖĖĒĢśĻ│Ā 242Ļ░£ņØś ņĢäļ»ĖļģĖņé░ņØä ņĢöĒśĖĒÖö(coding)ĒĢśļŖö Ļ▓āņØä ĒÖĢņØĖĒ¢łļŗż. ļČäļ”¼ņŻ╝ SHņÖĆ KHņØś CPņŚ╝ĻĖ░ ļ░Å ņĢäļ»ĖļģĖņé░ ņä£ņŚ┤ ņāüļÅÖņä▒ņØĆ Ļ░üĻ░ü 90.4ņÖĆ 95.5%ņśĆļŗż. ĻĖ░ņĪ┤ņŚÉ ļČäļ”¼ļÉ£ KNS1ņØä ĒżĒĢ©ĒĢ£ ĻĄŁļé┤ ļČäļ”¼ņŻ╝ 3ņóģņØś CP ņĢäļ»ĖļģĖņé░ ņāüļÅÖņä▒ņØĆ 95.5-99.8% (Table 2)ļĪ£, clustal X2 ņĀĢļĀ¼ Ļ▓░Ļ│╝ 10Ļ││(4, 20, 22, 31, 40, 67, 72, 74, 112, 158ļ▓łņ¦Ė ņä£ņŚ┤)ņŚÉņä£ ņĢäļ»ĖļģĖņé░ņØ┤ ņ╣śĒÖśļÉśņ¢┤ ņ׳ņŚłļŗż(Fig. 1). KNS1ļŖö SHņÖĆ Ļ░ÖņØĆ ĒÆłņóģ(ņäżĒ¢ź)ņŚÉņä£ ļČäļ”¼ļÉśņŚłņØīņŚÉļÅä KHņÖĆ ļŹö ļåÆņØĆ ņāüļÅÖņä▒(99.8%, Table 2)ņØä ļ│┤ņśĆņ£╝ļ®░ ļæÉ ļČäļ”¼ņŻ╝Ļ░ä ņ╣śĒÖśņĢäļ»ĖļģĖņé░ņØĆ 2Ļ││(31, 40ļ▓łņ¦Ė)ņØ┤ņŚłļŗż(Fig. 1). ĒĢ£ĒÄĖ, potexvirus ņåŹ ļ░öņØ┤ļ¤¼ņŖżņØś CPņŚÉņä£ Ļ│ĄĒåĄņĀüņ£╝ļĪ£ ļ│┤ņĪ┤ļÉśņ¢┤ ņ׳ļŗżĻ│Ā ņĢīļĀżņ¦ä ņŚ╝ĻĖ░ņä£ņŚ┤(consensus sequences)ņØĖ C ļ¦Éļŗ©ņØś FAAFDFFDGV (168-174aa residue)ņä£ņŚ┤ņØä SMYEV ĻĄŁļé┤ ļČäļ”¼ņŻ╝ņŚÉņä£ļÅä ĒÖĢņØĖ ĒĢĀ ņłś ņ׳ņŚłļŗż(Fig. 1, Bancroft ļō▒, 1991).
Table┬Ā2
Average percent nucleotide sequence and amino acid identities (number in bracket) between the three Korean isolates and the five phylogenetic groups of SMYEV
Fig.┬Ā1
Alignment of the complete amino acid (aa) sequences of the coat protein of 3 SMYEV Korean isolates. Sequences were aligned by ClustalX2. Consensus aa sequences are located line below. Only those residues differing from consensus are shown for the isolates. Region underlined is the most conserved motifs (FAAFDFF at residue 168-174) shared between SMYEV isolates.

Ļ│äĒåĄņ£ĀņĀäĒĢÖņĀü ņ£ĀņŚ░Ļ┤ĆĻ│ä ļČäņäØ
SMYEVņØś Ļ│äĒåĄļČäņäØņŚÉ ļīĆĒĢ£ ņŚ░ĻĄ¼ļŖö 2004ļģäņŚÉ ThompsonĻ│╝ JelkmannņŚÉ ņØśĒĢ┤ ņ▓śņØī ļ│┤Ļ│Ā ļÉśņŚłļŖöļŹ░ 24Ļ░£ ļČäļ”¼ņŻ╝ņŚÉ ļīĆĒĢ£ ļČäņäØ Ļ▓░Ļ│╝ 3Ļ░£ņØś subgroup (I, II, III)ņ£╝ļĪ£ ļČäļźśļÉśļ®░, ļČäļ”¼ļÉ£ ĻĖ░ņŻ╝ņÖĆ ņ¦ĆņŚŁņŚÉ ļö░ļØ╝ D74 (subgroup I), 9Redland (subgroup II), MY18 (subgroup III)ņØä ļīĆĒæ£ ļ│ĆņØ┤ņŻ╝(Type strain)ļź╝ ņ¦ĆņĀĢĒĢśņśĆļŗż(ThompsonĻ│╝ Jelkmann, 2004). ĻĘĖļ¤¼ļéś ņØ┤Ēøä Bhagwat ļō▒(2015)Ļ│╝ Torrico ļō▒(2016)ņØ┤ Ļ░üĻ░ü ņ║Éļéśļŗż ļČäļ”¼ņŻ╝ņÖĆ ņĢäļź┤ĒŚ©Ēŗ░ļéś ļČäļ”¼ņŻ╝ļź╝ ņČöĻ░ĆĒĢśņŚ¼ Ļ░üĻ░ü Ļ│äĒåĄļČäņäØņØä ĒĢ£ Ļ▓░Ļ│╝, 5Ļ░£(I, II, III, IV, V)ņÖĆ 4Ļ░£(I, II, III, IV)ņØś subgroupņØ┤ ņāłļĪŁĻ▓ī ĒśĢņä▒ļÉ£ phylogenetic treeļź╝ ņĀ£ņŗ£ĒĢ£ ļ░ö ņ׳ļŗż(Bhagwat ļō▒, 2015; Torrico ļō▒, 2016). ļ│Ė ļģ╝ļ¼ĖņŚÉņä£ļŖö ņāłļĪ£ ļ│┤Ļ│ĀĒĢśļŖö 2Ļ░£ņØś ĻĄŁļé┤ ļČäļ”¼ņŻ╝ņÖĆ Ēśäņ×¼ NCBIņŚÉ ļō▒ļĪØļÉ£ SMYEV ņÖĖĒö╝ļŗ©ļ░▒ņ¦łņŚÉ ļīĆĒĢ£ 46Ļ░£ ļČäļ”¼ņŻ╝ņØś ņŚ╝ĻĖ░ņä£ņŚ┤ ņĀĢļ│┤ļź╝ ļ¬©ļæÉ ņłśņ¦æĒĢśņŚ¼ ņ┤Ø 48Ļ░£ņŚÉ ļīĆĒĢ£ Ļ│äĒåĄĒĢÖņĀü ņ£ĀņŚ░Ļ┤ĆĻ│äļź╝ ļČäņäØĒĢśņśĆļŗż. Neighbor-joining ļČäņäØļ░®ņŗØņŚÉ ņØśĒĢ£ phylogenetic tree ņ×æņä▒ Ļ▓░Ļ│╝ 5Ļ░£ņØś subgroupņØ┤ ĒśĢņä▒ļÉ£ treeļź╝ ņ¢╗ņŚłņ£╝ļ®░ ņØ┤ 5Ļ░£ groupņØĆ Ēü¼Ļ▓ī 3Ļ░£ņØś subgroup (I, II, IV)ņ£╝ļĪ£ ļČäļ”¼ļÉśļŖö group A ņÖĆ 2Ļ░£ņØś subgroup (III, V)ņ£╝ļĪ£ ļéśļēśļŖö group BļĪ£ ĒśĢņä▒ļÉ£ treeņØś ĒśĢĒā£ņśĆņ£╝ļ®░ Ļ░ü group AņÖĆ BņØś bootstrap valueļŖö 71%ņÖĆ 82%ņśĆļŗż(Fig. 2). ĻĄŁļé┤ ļČäļ”¼ņŻ╝ KHņÖĆ KNS1ņØĆ subgroup IņŚÉ ĒżĒĢ©ļÉ£ ļ░śļ®┤, SHļŖö subgroup IVļĪ£ ļČäļźśļÉśņ¢┤ ĻĄŁļé┤ ļČäļ”¼ņŻ╝ Ļ░äņŚÉļÅä ļ│ĆņØ┤Ļ░Ć ņ׳ņØīņØä ņĢī ņłś ņ׳ņŚłļŗż(Fig. 2). Subgroup IņØĆ 48Ļ░£ ļČäļ”¼ņŻ╝ ņżæ 24Ļ░£ ļČäļ”¼ņŻ╝Ļ░Ć ĒżĒĢ©ļÉśņŚłņ£╝ļ®░ ĻĄŁļé┤ ļČäļ”¼ņŻ╝ 2Ļ░£, ņ£Āļ¤Į ļČäļ”¼ņŻ╝ 12Ļ░£(ļÅģņØ╝ 8Ļ░£, ļ▓©ĻĖ░ņŚÉ 1Ļ░£, ņØ┤Ēāłļ”¼ņĢä 1Ļ░£, ņ▓┤ņĮö 2Ļ░£), ņĢäļź┤ĒŚ©Ēŗ░ļéś ļČäļ”¼ņŻ╝ 4Ļ░£, ļ»ĖĻĄŁ ļČäļ”¼ņŻ╝ 2Ļ░£, ņżæĻĄŁ ļČäļ”¼ņŻ╝ 2Ļ░£Ļ░Ć ĒżĒĢ©ļÉśņŚłļŗż. ļŗżņ¢æĒĢ£ ņ¦ĆņŚŁņØś ļČäļ”¼ņŻ╝Ļ░Ć ĒĢ£ ļČäļźśĻĄ░ņŚÉ ĒżĒĢ©ļÉśļŖö Ļ▓āņ£╝ļĪ£ ļ│┤ņĢä ņ¦ĆņŚŁĻ░ä Ļ│äĒåĄņĀü ņ£ĀņØśņä▒ņØĆ ņŚåļŖö Ļ▓āņ£╝ļĪ£ ņé¼ļŻīļÉ£ļŗż. Subgroup IņØś 24Ļ░£ ļČäļ”¼ņŻ╝ļōż Ļ░äņØś ņŚ╝ĻĖ░ņä£ņŚ┤ ņāüļÅÖņä▒ņØĆ 96.3-100%ņśĆņ£╝ļ®░ KHĻ│╝ KNS1 ĻĄŁļé┤ ļČäļ”¼ņŻ╝ļŖö ļ»ĖĻĄŁļČäļ”¼ņŻ╝ņØĖ WSU1988ņÖĆ 98.6%ļĪ£ Ļ░Ćņן Ļ░ĆĻ╣īņÜ┤ ņ£ĀņŚ░Ļ┤ĆĻ│äļź╝ ļ│┤ņśĆļŗż. Subgroup IIļŖö 8Ļ░£ ļČäļ”¼ņŻ╝(ĒśĖņŻ╝ 1Ļ░£, ņĢäļź┤ĒŚ©Ēŗ░ļéś 3Ļ░£, ņ╣ĀļĀł4Ļ░£), Subgroup IIIļŖö 5Ļ░£ ļČäļ”¼ņŻ╝(ļ»ĖĻĄŁ 1Ļ░£, ņĢäļź┤ĒŚ©Ēŗ░ļéś 1Ļ░£, ņ╣ĀļĀł 3Ļ░£)Ļ░Ć ĒżĒĢ©ļÉśņŚłĻ│Ā Ļ░ü ļČäļźśĻĄ░ ļé┤ ļČäļ”¼ņŻ╝ Ļ░ä ņŚ╝ĻĖ░ņä£ņŚ┤ ņāüļÅÖņä▒ņØĆ Ļ░üĻ░ü 94.9-98.9%ņÖĆ 97.9-99%ņśĆļŗż. ĒĢ£ĒÄĖ, ļśÉ ļŗżļźĖ ĻĄŁļé┤ ļČäļ”¼ņŻ╝ SHļŖö ņ║Éļéśļŗż ļČäļ”¼ņŻ╝ 3Ļ░£ņÖĆ ĒĢ©Ļ╗ś subgroup IVļĪ£ ļČäļźśļÉśņŚłļŖöļŹ░ ļČäļ”¼ņŻ╝Ļ░ä ņŚ╝ĻĖ░ņä£ņŚ┤ ņāüļÅÖņä▒ņØĆ 95.6-100% ņśĆņ£╝ļ®░ ĻĘĖ ņżæ SHņÖĆ AB5-2ļŖö ņŚ╝ĻĖ░ņä£ņŚ┤ņØ┤ 100% ņØ╝ņ╣śĒ¢łļŗż. Subgroup VļŖö 7Ļ░£ ļČäļ”¼ņŻ╝(ņ║Éļéśļŗż 2Ļ░£, ņżæĻĄŁ 2Ļ░£, ņĢäļź┤ĒŚ©Ēŗ░ļéś 3Ļ░£)Ļ░Ć ĒżĒĢ©ļÉśņŚłņ£╝ļ®░ ņŚ╝ĻĖ░ņä£ņŚ┤ ņāüļÅÖņä▒ņØĆ 83.3~99.2%ļĪ£ Ļ░Ćņן ļé«ņØĆ ņāüļÅÖņä▒ņØä ļ│┤ņśĆņ£╝ļéś bootstrap valueļŖö 97%ļĪ£ ļåÆĻ▓ī ļéśĒāĆļé¼ļŗż.
Fig.┬Ā2
Phylogenetic analysis of the complete nucleotide sequences of the coat protein of different SMYEV isolates (Table 1). Three Korean isolates are marked with arrows. The Potato virus X (PVX; GenBank accession No. NC011620) was included as an outgroup. The phylogenetic trees were reconstructed by the NJ method applying KimuraŌĆÖs two-parameter method. The numbers on the branches indicate bootstrap percentages based on 1000 replications (only values >60% are shown). The scale bar represents the number of substitutions per base.
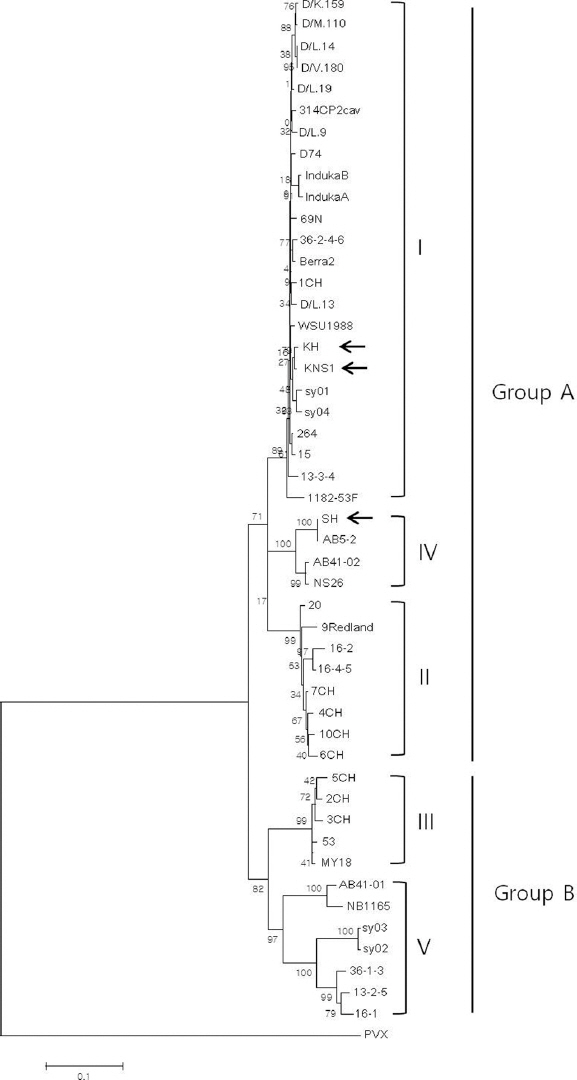
5Ļ░£ subgroupĻ░ä ņ£ĀņĀäņĀü ĻĘ╝ņŚ░Ļ┤ĆĻ│äļź╝ ļ│┤ĻĖ░ ņ£äĒĢ┤ Ļ░ü subgroupļ│ä 5Ļ░£ ļīĆĒæ£ ļ│ĆņØ┤ņŻ╝(I, D74; II, 9Redland; III, MY18; IV, AB41-02; V, AB41-01)ļź╝ ņäĀļ░£ĒĢśĻ│Ā ĻĄŁļé┤ ļČäļ”¼ņŻ╝ 3Ļ░£ņÖĆ ĒĢ©Ļ╗ś ņŚ╝ĻĖ░ņÖĆ ņĢäļ»ĖļģĖņé░ņä£ņŚ┤ ņāüļÅÖņä▒ņØä ļČäņäØĒĢśņśĆļŗż(Table 2). Group AņŚÉ ĒżĒĢ©ļÉ£ subgroup IĻ│╝ IVļŖö ņŚ╝ĻĖ░ņä£ņŚ┤(91.8%)Ļ│╝ ņĢäļ»ĖļģĖņé░ņä£ņŚ┤(96.7%) ļ¬©ļæÉ ņä£ļĪ£ Ļ░Ćņן ļåÆņØĆ ņāüļÅÖņä▒ņØä ļ│┤ņśĆņ£╝ļ®░, subgroup IIļŖö IĻ│╝ ņŚ╝ĻĖ░(91.1%)ņÖĆ ņĢäļ»ĖļģĖņé░(96.3%) ņä£ņŚ┤ ņāüļÅÖņä▒ņØ┤ Ļ░Ćņן ļåÆņĢśļŗż. Group BņŚÉ ĒżĒĢ©ļÉ£ subgroup IIIņØś Ļ▓ĮņÜ░, ņŚ╝ĻĖ░ņä£ņŚ┤ ņāüļÅÖņä▒ņØĆ 88.5%ļĪ£ IVņÖĆ Ļ░Ćņן ļåÆņØĆ ļ░śļ®┤ ņĢäļ»ĖļģĖņé░ ņä£ņŚ┤ņØĆ 95.1%ļĪ£ I, II ļÅÖņØ╝ĒĢśĻ▓ī Ļ░Ćņן ļåÆņĢśļŗż. Subgroup VļŖö ņŚ╝ĻĖ░(85.7%), ņĢäļ»ĖļģĖņé░(94.7%) ņä£ņŚ┤ ļ¬©ļæÉ IIIņÖĆ Ļ░Ćņן ļåÆņØĆ ņāüļÅÖņä▒ņØä ļ│┤ņśĆļŗż(Table 2). Phylogenetic treeņÖĆ ņŚ╝ĻĖ░ņä£ņŚ┤ ņāüļÅÖņä▒ ļČäņäØ Ļ▓░Ļ│╝, ņāłļĪ£ņÜ┤ 2Ļ░£ subgroupņżæ IVļŖö subgroup IĻ│╝ IIņŚÉņä£ ĒīīņāØļÉ£ ļ│ĆņØ┤ ļČäļźśĻĄ░ņØ┤ļ®░ subgroup VļŖö subgroup IIIņŚÉņä£ ĒīīņāØļÉ£ ļ│ĆņØ┤ ļČäļźśĻĄ░ņØĖ Ļ▓āņ£╝ļĪ£ ņČöņĖĪļÉ£ļŗż. ņāłļĪ£ņÜ┤ ļ│ĆņØ┤ ļČäļźśĻĄ░ ļ░£ņāØņŚÉ ļīĆĒĢ£ phylogenetic tree ļČäņäØņØĆ SMYEVņØś ņ¦äĒÖö ļ░Å ļŗżņ¢æņä▒ņØä ņØ┤ĒĢ┤ĒĢśļŖö ĻĖ░ņ┤łņ×ÉļŻīĻ░Ć ļÉĀ Ļ▓āņ£╝ļĪ£ ņé¼ļŻīļÉ£ļŗż.
ņ£ĀņĀäņĀü ļŗżņ¢æņä▒ ļČäņäØ
SMYEVņØś ņāłļĪ£ņÜ┤ ļ│ĆņØ┤ņ¦æļŗ©ņØ┤ ĒśĢņä▒ļÉśļŖö ņøÉņØĖņØĆ ļ░öņØ┤ļ¤¼ņŖżĻ░Ć ņ£ĀņĀäņĀüņ£╝ļĪ£ ļŹö ļŗżņ¢æĒĢśĻ▓ī ņ¦äĒÖöļÉśĻ│Ā ņ׳ņØīņØä ņØśļ»ĖĒĢ£ļŗż. ĒŖ╣Ē׳ RNA Ļ▓īļåł ļ░öņØ┤ļ¤¼ņŖżļŖö ļ│ĆņØ┤(mutation)ņÖĆ ņ×¼ņĪ░ĒĢ®(recombination)ņØś ļ╣äņ£©ņØ┤ ļåÆņĢä ņ£ĀņĀäņ×É ļŗżņ¢æņä▒ņØś ļ╣łļÅäĻ░Ć ļåÆņØĆ Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż(Garcia-Arenal ļō▒, 2001; RobinsonĻ│╝ Murray, 2013). SMYEVņØś Ļ▓ĮņÜ░ phylogenetic tree ļČäņäØ Ļ▓░Ļ│╝ ņāłļĪ£ņÜ┤ ļ│ĆņØ┤ņŻ╝ļōżņØ┤ ļ░£ņāØļÉśĻ│Ā ņ׳ņØīņØ┤ ņŗ£ņé¼ļÉśņ¢┤ Ļ░ü subgroupņŚÉ ļīĆĒĢ£ ņ£ĀņĀäņ×É ļŗżņ¢æņä▒ ļČäņäØņØä ņŗżņŗ£ĒĢśņśĆļŗż. Ļ░ü subgroupņØś ļČäļ”¼ņŻ╝ Ļ░ä Ēś╣ņØĆ ņ¦æļŗ© Ļ░äņŚÉ ļéśĒāĆļéśļŖö ņŚ╝ĻĖ░ņä£ņŚ┤ ļŗżņ¢æņä▒(nucleotide divergence)Ļ│╝ subgroupĻ░ä ļŗżĒśĢņä▒(polymorphism)ņØś ņÜöņØĖļōżņØä ļČäņäØĒ¢łļŗż. Ļ░ü subgroup (I-V)ņØś ņŚ╝ĻĖ░ņä£ņŚ┤ ļŗżņ¢æņä▒ņØĆ Ļ░üĻ░ü 0.01864(I), 0.02660(II), 0.01838(III), 0.02812(IV), 0.11584(V)ļĪ£ ļéśĒāĆļéś subgroup VņØś ļŗżņ¢æņä▒ņØ┤ Ļ░Ćņן ļåÆņØīņØä ņĢī ņłś ņ׳ņŚłļŗż. ļ¬©ļōĀ ļČäļ”¼ņŻ╝ņŚÉ ļīĆĒĢ£ ņŚ╝ĻĖ░ņä£ņŚ┤ ļŗżņ¢æņä▒ ņłśņ╣śļŖö 0.0993ņØ┤ņśĆļŗż. SubgroupĻ░ä ņŚ╝ĻĖ░ņä£ņŚ┤ ļŗżņ¢æņä▒ņØĆ 0.08634-0.18004ļĪ£ subgroup IĻ│╝ IVĻ░Ć Ļ░Ćņן ļé«Ļ│Ā, subgroup IIņÖĆ VĻ░Ć Ļ░Ćņן ļåÆņĢśļŗż(Table 3). ļŗżņ¢æņä▒ ņ¦ĆņłśļŖö ņ£ĀņĀäĻ▒░ļ”¼ļź╝ ļéśĒāĆļé┤ļŖö Ļ▓āņ£╝ļĪ£ ņŚ╝ĻĖ░ņä£ņŚ┤ņØś ņ£Āņé¼ņä▒ņŚÉ ĻĘ╝Ļ▒░ĒĢśļ»ĆļĪ£ ņłśņ╣śĻ░Ć ļé«ņØäņłśļĪØ ņ£ĀņĀäņĀü ņ£ĀņŚ░Ļ┤ĆĻ│äĻ░Ć ļåÆņØīņØä ņØśļ»ĖĒĢ£ļŗż(Rogers ļō▒, 2012). Subgroup Ļ░ä ļŗżĒśĢņä▒ ļČäņäØĻ▓░Ļ│╝ļŖö Table 4ņŚÉ ņĀĢļ”¼ĒĢśņśĆļŗż. ņ£ĀņĀäņ×É ļŗżĒśĢņä▒ ļČäņäØņŚÉ Ļ░Ćņן Ēü░ ņÜöņØĖņØ┤ ļÉśļŖö polymorphic siteņÖĆ parsimony informative siteņØś ļ╣äņ£© ļČäņäØ Ļ▓░Ļ│╝, Ļ░ü subgroup ļ│ä ļ╣äņ£©ņØĆ I: 8.2%ņÖĆ 5.2%; II: 8.9%ņÖĆ 2.1%; III: 4.3%ņÖĆ 0.7%; IV: 4.4%ņÖĆ 3.7%; V: 24.6%ņÖĆ 16.6%ļĪ£ subgroup VĻ░Ć Ļ░Ćņן ļåÆņØĆ ļŗżĒśĢņä▒ņØä Ļ░Ćņ¦ĆĻ│Ā ņ׳ļŖö Ļ▓āņØä ĒÖĢņØĖĒĢĀ ņłś ņ׳ņŚłļŗż. ĒĢ£ĒÄĖ ļ¬©ļōĀ ļČäņäØ ņŚ╝ĻĖ░ņä£ņŚ┤ņŚÉņä£, ņĮöļÅł(codon)ņŚÉļŖö ņśüĒ¢źņØä ņŻ╝ņ¦Ć ņĢŖņ£╝ļ®┤ņä£ ņ£ĀņĀäņ×Éļź╝ ļ│ĆņØ┤ņŗ£ĒéżļŖö ļÅÖņØśņ╣śĒÖś(synonymous substitution)ņØś ņłśĻ░Ć ļ╣äļÅÖņØśņ╣śĒÖś(synonymous substitution)ņŚÉ ļ╣äĒĢ┤ ļŹö ļåÆĻ▓ī ļéśĒāĆļé¼ļŖöļŹ░ ĒŖ╣Ē׳ subgroup VņØś Ļ▓ĮņÜ░ ļÅÖņØśņ╣śĒÖśņØś ņłśĻ░Ć 141ļĪ£ ļŗżļźĖ subgroupļ│┤ļŗż Ēø©ņö¼ ļåÆņĢśļŗż(Table 4). ĻĄŁļé┤ ļČäļ”¼ņŻ╝ņØś Ļ▓ĮņÜ░ subgroup IĻ│╝ IVņŚÉļ¦ī ĒżĒĢ©ļÉśņ¦Ćļ¦ī ņāłļĪ£ņÜ┤ ļ│ĆņØ┤ņŻ╝Ļ░Ć ņČ£ĒśäļÉśļŖöņ¦Ć ņ¦ĆņåŹņĀüņØĖ ņŚ░ĻĄ¼Ļ░Ć ĒĢäņÜöĒĢĀ Ļ▓āņ£╝ļĪ£ ņŚ¼Ļ▓©ņ¦äļŗż. SMYEVņØś ņ£ĀņĀäņ×É ļŗżņ¢æņä▒ņŚÉ ļö░ļźĖ ļ│ĆņØ┤ņŻ╝ļōżņØś ļ░öņØ┤ļ¤¼ņŖż ļ│æņøÉņä▒ ļ░Å ĻĖ░ņŻ╝ļ▓öņ£ä, ļöĖĻĖ░ ĒÆłņóģĻ░ä ņĀĆĒĢŁņä▒ ņ░©ņØ┤ ļō▒ņØĆ ņĢ×ņ£╝ļĪ£ ļŹö ņŚ░ĻĄ¼Ļ░Ć ņ¦äĒ¢ēļÉśņ¢┤ņĢ╝ ĒĢĀ Ļ▓āņ£╝ļĪ£ ņé¼ļŻīļÉ£ļŗż.
Table┬Ā3
Nucleotide divergence of CP gene within and between subgroups of all SMYEV isolates
Table┬Ā4
Information about the polymorphism of SMYEV isolate subgroups
ņÜö ņĢĮ
ĻĄŁļé┤ņ£Īņä▒ļöĖĻĖ░ ĒÆłņóģņØĖ ņäżĒ¢źĻ│╝ Ļ░ÉĒÖŹņŚÉņä£ ļöĖĻĖ░ļłä ļźĖņśżĻ░łļ░öņØ┤ļ¤¼ņŖżņØś ĻĄŁļé┤ ļČäļ”¼ņŻ╝ 2ņóģņØä ļČäļ”¼ĒĢśĻ│Ā ņÖĖĒö╝ļŗ©ļ░▒ņ¦ł ņĀäņ▓┤ ņŚ╝ĻĖ░ņä£ņŚ┤ņØä Ļ▓░ņĀĢĒĢśĻ│Ā ļČäņäØĒĢśņśĆļŗż. ĻĄŁļé┤ ļČäļ”¼ņŻ╝ SHņÖĆ KHņØś ņÖĖĒö╝ļŗ©ļ░▒ņ¦ł ņŚ╝ĻĖ░ņÖĆ ņĢäļ»ĖļģĖņé░ ņāüļÅÖņä▒ņØĆ Ļ░üĻ░ü 90.4%ņÖĆ 95.5% ņśĆļŗż. ĻĖ░ņĪ┤ņŚÉ ĻĄŁļé┤ņŚÉņä£ ļ│┤Ļ│ĀļÉ£ KNS1ļČäļ”¼ņŻ╝ņÖĆ GenBankņŚÉ ļō▒ļĪØļÉ£ 45Ļ░£ņØś ļŗżļźĖļéśļØ╝ ļČäļ”¼ņŻ╝ ņÖĖĒö╝ļŗ©ļ░▒ņ¦ł ņŚ╝ĻĖ░ņä£ņŚ┤ņØä ļ¬©ļæÉ ņłśņ¦æĒĢśņŚ¼ ņ┤Ø 48Ļ░£ SMYEV ņÖĖĒö╝ļŗ©ļ░▒ņ¦łņŚÉ ļīĆĒĢ£ Ļ│äĒåĄĒĢÖņĀü ņ£ĀņŚ░Ļ┤ĆĻ│äļź╝ ļČäņäØĒĢĀ Ļ▓░Ļ│╝ ņ┤Ø 5Ļ░£ņØś subgroup (I-V)ņ£╝ļĪ£ ļČäļźśĻ░Ć ļÉśņŚłļŗż. ņØ┤ ņżæ subgroup IVĻ│╝ VĻ│╝ ņāłļĪ£ņÜ┤ ļ│ĆņØ┤ņ¦æļŗ©ņ£╝ļĪ£ ĻĄŁļé┤ļČäļ”¼ņŻ╝ļÅä KHņÖĆ KNS1ņØĆ subgroup IņŚÉ ĒżĒĢ©ļÉ£ ļ░śļ®┤, SHļŖö ņāłļĪ£ņÜ┤ subgroupņØĖ IVņŚÉ ĒżĒĢ©ļÉśņ¢┤ ĻĄŁļé┤ļČäļ”¼ņŻ╝Ļ░äņŚÉļÅä Ļ│äĒåĄņØ┤ ļŗżļźĖ Ļ▓āņØä ņČöņĖĪĒĢĀ ņłś ņ׳ņŚłļŗż. ņ£ĀņĀäņĀü ļŗżņ¢æņä▒ ļČäņäØĻ▓░Ļ│╝ SMYEVņØś ņāłļĪ£ņÜ┤ subgroupņØś ļŗżņ¢æņä▒ņØ┤ ļŹöņÜ▒ ļåÆņØĆ Ļ▓āņ£╝ļĪ£ ļéśĒāĆļéś SMYEVĻ░Ć ņ£ĀņĀäņĀüņ£╝ļĪ£ ņ¦äĒÖöļź╝ ĒĢśĻ│Ā ņ׳ņØīņØä ņĢī ņłś ņ׳ņŚłļŗż. ņØ┤ ļģ╝ļ¼ĖņØĆ ĻĄŁļé┤ SMYEV ļČäļ”¼ņŻ╝ņŚÉ ļīĆĒĢ£ ļČäņ×ÉņĀü ĒŖ╣ņä▒ņŚÉ ļīĆĒĢ£ ņ▓½ ļ│┤Ļ│ĀņØ┤ļŗż.
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






