


Plant possess elegant and effective survival systems against abiotic stress and biotic invasion (Zhu, 2001). Under drought conditions, the level of the plant hormone, abscisic acid (ABA) increases, triggering adaptive responses that are essential for the survival of plants (Zhu, 2001). One effect of the enhanced ABA level is stomatal closure that minimizes water loss through transpiration (Leung and Giraudat, 1998). Consequently, because of the problems associated with the regulation of stomatal aperture, plants with defects in ABAbiosynthesis (aba mutants) and ABA-responses (i.e., ABAinsensitive abi mutants) are susceptible to drought stress (Leung and Giraudat, 1998; Schroeder et al., 2001). Another plant growth regulator, jasmonate (Evans, 2003; Suhita et al., 2004), also stimulates stomatal closure under drought conditions (Creelman and Mullet, 1997).
In plants, drought tolerance can be induced upon colonization by certain microbes. Pseudomonas chlororaphis O6, a plant probiotic microbe (Kim and Anderson, 2018), elicits both induced systemic resistance (ISR) to pathogens and induced systemic tolerance (IST) to environmental stresses (Kim et al., 2011; Yang et al., 2009, 2017). Induced tolerance to drought in plants also occurs with colonization by several Gram-positive Bacillus strains (Ryu et al., 2004; Timmusk and Wagner, 1999; Zhang et al., 2010), other Gram-negative bacterial isolates (Mayak et al., 2004), and fungi such as Trichoderma harziarum (Bae et al., 2008; Shukla et al., 2015). The volatile 2R,3R-butanediol is a metabolite produced by the plant-associated bacteria strains that causes stomatal closure and induces the expression of genes promoting drought tolerance (Kim et al., 2012). Butanediol-induced drought tolerance in Arabidopsis involves activation of salicylic acid (SA), jasmonic acid, and ethylene-dependent pathways (Cho et al., 2008).
Recent reviews indicate that there is an array of different types of microbial determinants inducing systemic tolerance in plants against biotic and/or abiotic stresses (Bakker et al., 2013; Cheng et al., 2017; Kim and Anderson, 2018). Some of the determinants have been identified by examining mutants in root-colonizing beneficial microbes impaired in their abilities to cause IST (Cheng et al., 2017; Han et al., 2006; Kim et al., 2007). The active microbial structures include such secreted metabolites (Kim and Anderson, 2018) as antibiotics, siderophores, and organic volatiles (e.g. butanediol) and bacterial surface structures, the lipopolysaccharides and flagella proteins (Melotto et al., 2006; Underwood et al., 2007). These findings illustrate that a single microbe may produce several factors that will cause IST.
We observed that induced drought tolerance by P. chlororaphis O6 correlates with decreased stomatal aperture size on the leaf in plants with roots colonized by this microbe (Cho et al., 2008). In this study, we extend the knowledge of microbial products inducing tolerance to drought to include the extracellular polymeric substances (EPS) from P. chlororaphis O6. The potential of EPS to induce drought tolerance was studied by determining whether they caused stomatal closure in the epidermal peels. At the whole plant level, the extent of wilting upon withholding water was assessed by visual comparison between plants with and without EPS treatments. The extent of drought was also characterized by measuring the chlorophyll fluorescence in the leaf tissues (Woo et al., 2008). Drought causes changes in the photosynthetic potential of plants that can be detected by measuring the Fv/Fm ratio (Woo et al., 2008).
Isolation of EPS from P. chlororaphis O6
Cells from a single colony of P. chlororaphis O6, grown for 2 d on Luria-Broth (LB, Difco Inc., Detroit, MI, USA) agar plates, were inoculated into 500 ml of LB broth and incubated at room temperature for 4 d with agitation at 250 rpm. The EPS of P. chlororaphis O6 were isolated using a modified ethanol precipitation method as described previously (Hung et al., 2005). The bacterial cells were removed by centrifugation for 10 min, and 30 Anson unit protease (0.5 mg/l, Quiagen Inc., Valencia, CA, USA) was added to the supernatant and incubated at 37┬░C for 12 h with agitation at 70 rpm. Ethanol was added to the supernatant to produce a 1:4 (v/v) mixture of the supernatant:ethanol and stirred at 4┬░C for 14 h. The precipitate was removed by filtration through a 0.2 ╬╝m Millipore membrane. The precipitate was dissolved in sterile water and mixed with ethanol (1:4 ratio) at 4┬░C for 14 h. The EPS were pelleted by centrifugation for 15,000├Śg at 4┬░C for 30 min. The pellet was dried at room temperature for 5 hand the mass measured before dissolving in sterile distilled water to produce solutions with known mass/ml. The final EPS preparation was streaked on LB agar plate to confirm no contamination by any culturable microbes.
Data were analyzed by ANOVA (p<0.01), using SPSS (version 23, SPSS Inc., Chicago, IL, USA) and if the value of the F was significant, differences between each measurement were further elucidated through DuncanŌĆÖs multiple range test (p<0.01).
Examination of stomatal opening on epidermal peels
To determine the direct effects of EPS on stomatal opening, plants were grown in well plates for 4 weeks. Arabidopsis thaliana Col-O seeds were surfaced-sterilized with 0.3% sodium hypochlorite, followed by extensive washing with sterile water. Seeds were transferred to well plates (12-well microtiter plates; SPL Inc., Seoul, South Korea) that contained half-strength Murashige and Skoog salt (MS, Sigma-Aldrich, St. Louis, MO, USA) medium, 0.3% Phytagel and 3% sucrose (pH 5.7) (Cho et al., 2008, 2012). The plates were sealed with Parafilm to prevent drying and no additional water was added during their growth. To maximize stomatal opening, the plants were maintained under light for at least 3 h before excising the epidermal peels. The peels were transferred on to glass slides. The cuticle side was treated with water (a negative control), defined dilutions of the EPS solution (10, 100, and 1000 ╬╝g/ml, the test treatments), or a suspension of P. chlororaphis O6 cells (a positive control). Another positive control for stomatal closure was treatment with 100 ╬╝M ABA (Sigma-Aldrich). Two hours after incubation, the stomatal apertures were observed by light microscopy (Leica M165 FC, Leica Microsystems, Wetzlar, Germany). The size of the openings was assessed using the image acquisition software, Image-Pro (version 2.32 for Windows, Media Cybermetics, Rockville, MD, USA). At least 100 stomata/plants were counted, with five plants per treatment being used in each experimental replicate.
The treatment of leaf epidermal peels with ABA and with the suspension of intact P. chlororaphis O6 cells decreased the size of stomatal aperture relative to stomata treated exclusively with water (Fig. 1A). These findings were expected based on the previous findings (Cho et al., 2008). Stomatal closure also was observed with treatment of 100 ╬╝g/ml EPS solution (Fig. 1A-C). The extent of closure was dose-dependent (Fig. 1B); when doses of 10, 100, and 1000 ╬╝g/ml were used, the 100 ╬╝g/ml treatment was optimal.
Fig.┬Ā1
Stomatal closure in epidermal peels by treatments with Pseudomonas chlororaphis O6 cells, its EPS and ABA. The treatments for 2 h were (A): water; 100 ╬╝M abscisic acid (ABA), or P. chlororaphis O6 cells (1├Ś108 cfu/ml) applied to the peel in 10 ╬╝l aliquots and EPS (100 ╬╝g/ml) in (B) water or P. chlororaphis O6 EPS at defined concentrations. The width of the stomatal apertures was measured from light microscope images using the software Image-Pro C. (C) The % closed stomates with treatments of water, EPS (100 ╬╝g/ml) or P. chlororaphis O6 cells (1├Ś108 cfu/ml). At least 100 stomata per plant were counted, with five plants per treatment used in each experimental replicate. Different letters indicate significant differences between treated and control samples based on DuncanŌĆÖs multiple range test at p<0.01.

Plant wilting bioassay
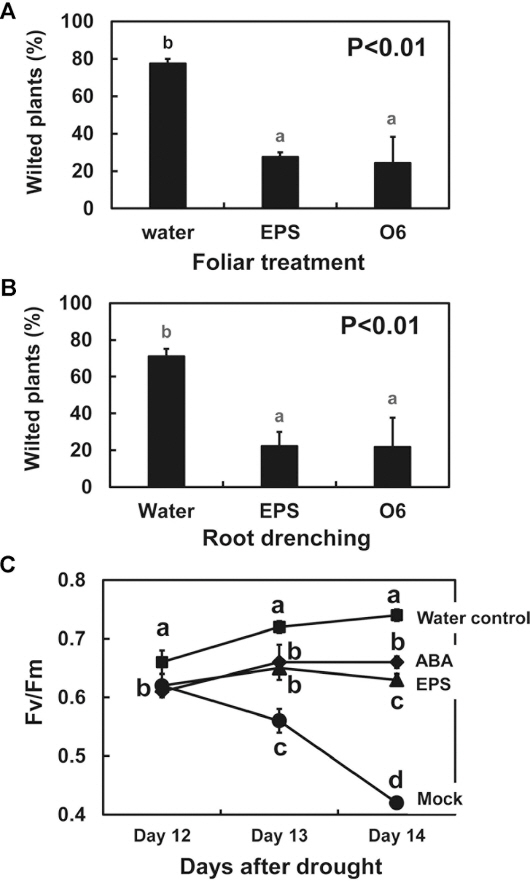
To determine whether the treatments that caused the narrowing of stomatal openings corresponded with drought tolerance, the degree of wilting was assessed visually using A. thaliana seedlings grown in the well plates as described previously (Cho et al., 2008). The surface-sterilized seeds were placed on Whatman # 1 paper and placed over the MS medium contained in the microtiter wells. The microtiter plates were sealed with parafilm to prevent drying. After two weeks of growth, the seedlings were treated with 10 ╬╝l/plant, water as a control, 10 ╬╝l per plant, P. chlororaphis O6 cell suspensions (1├Ś108 cfu/ml), or 10 ╬╝l per plant, 100 ╬╝g/ml EPS solution. After 3 d, seedlings on the filter papers were transferred to open, empty Petri dishes. The extent of wilting of the plants was assessed visually after 2 h. The experiment was repeated three times with 12 plants per treatment.
When A. thaliana seedlings were exposed to the air, prior treatments for 3 d with EPS, or with live P. chlororaphis O6 cells, resulted in plants that exhibited less visible wilting than the plants treated with water as a control (Fig. 2A). Thus, P. chlororaphis O6 cells and extracted EPS reduced the wilt symptoms in intact plants deprived of water.
Fig.┬Ā2
Protective effects of Pseudomonas chlororaphis O6 and its EPS against drought stress in Arabidopsis thaliana Col-0 seedlings grown in MS agar. The seedlings leaves (A) and roots (B) were treated with water (Water) as a control, P. chlororaphis O6 cells (O6, 1├Ś108 cfu/ml), or 100 ╬╝g/ml EPS. The extent of wilting of the plants deprived of water was assessed visually after 2 h. The study was performed three times with 12 plants per treatment. (C) Change in the Fv/Fm value of leaves of A. thaliana Col-0 plants raised in sterile soil mix. A subset of the plants had water withheld for 15 d whereas the control set had normal watering (water control). Treatments were water (mock), 100 ╬╝M ABA or EPS (100 ╬╝g/ml). The fluorescence measurements were recorded daily to compare the Fv/Fm values. The study was performed two times with 3 plants per treatment. The data presented are means and standard deviation from two studies. Different letters indicate significant differences between treated and control samples based on DuncanŌĆÖs multiple range test at p<0.01.
Quantitative assessment of drought survival using chlorophyll fluorescence
The changes in photosynthetic potential with drought were assessed with A. thaliana seedlings grown in sterile soil mix. The solid growth matrix was a sterile nursery soil mixture of Bio-Sangto, vermiculite, perlite (125 g/vessel, 3:1:1; Seminis Korea, Seoul, Korea) contained in sterile Magenta boxes (10 cm ├Ś 10 cm ├Ś 9 cm, Sigma-Aldrich). Five milliliters of water were added daily to the boxes. The seedlings were grown under a 16 h/8 h light/dark cycle with 40-W fluorescent lights (3,000 lx, 80 ╬╝mol photon m-2s-1). The temperature was maintained at 23┬▒2┬░C with a relative humidity of 50-60%.
The seedlings were grown for 3 weeks before treatment with 2 ml of sterile water as a negative control, or with equal volumes of 100 ╬╝M ABA as a positive control, or P. chlororaphis O6 EPS (100 ╬╝g/ml). A subset of the seedlings was drought-stressed by withholding water. After 12 d of drought stress, the chlorophyll fluorescence in the leaf tissues was measured using a Phyto-PAM (Heinz-Walz GmbH, Effeltrich, Germany) as described previously (Woo et al., 2008). The Fv/Fm was determined by the application of a single saturating pulse to dark-adapted leaves for 30 min and used as a quantitative measure of drought tolerance (Woo et al., 2008). The experiment was repeated twice with 3 plants per treatment.
In plants grown in pot cultures that were maintained with normal hydration, the Fv/Fm value for the leaves increased daily, indicating healthy photosynthesizing leaves (Fig. 2B). In plants that were subjected to drought stress by withholding water, the Fv/Fm value steadily declined with the number of days of stress due to the impaired functioning of photosystem II. However, plants that were subjected to water withholding when treated with either ABA or EPS had leaves that showed no reduction in the Fm/Fv values (Fig. 2B). This response was expected, because the partial closure of the stomata maintained function of photosystem II. Thus, treatments of the intact plants with the EPS from P. chlororaphis was as effective as ABA in protecting the plant from reduced photosynthetic efficacy under drought stress.
The present study indicated that the ethanol-precipitated EPS from P. chlororaphis O6 were effective in inducing protection against drought, as measured by limited wilting and by the maintenance of photosynthetic efficacy upon water withholding in intact Arabidopsis plants. Leaf tissues exposed to the EPS showed stomatal closure eliciting the same plant response as 2R, 3R-butanediol (Cho et al., 2008, 2013). Consequently, we demonstrated that EPS from P. chlororaphis O6 is another plant immune effector activating drought tolerance.
The EPS are part of the matrix that embeds bacterial cells in biofilms (Anderson and Kim, 2018). The composition of EPS likely includes different polysaccharides and extracellular DNA (Hung et al., 2005; Wingender et al., 1999). P. chlororaphis O6 forms patchy but layered and complex biofilms on roots as part of the surface colonization process (Bonebrake et al., 2018). Timmusk et al. (2015), based on their work with root-colonizing Gram-positive bacteria that active IST, concluded that the biofilm formation is required in induction of drought tolerance. Thus, the EPS produced in the root surface biofilms may be part of the microbial ŌĆ£microbe-associated molecular patternsŌĆØ (MAMPs) that activate plant immunity. Indeed, EPS from Bacillus cereus acts as a MAMP for stimulating plant disease resistance (Jiang et al., 2016). We are currently characterizing the components in the EPS from P. chlororaphis O6. An interesting question is whether alginate is present. Alginate present in the EPS from P. aeruginosa, xanthan from Xanthomonas campestris, and amylovoran from Erwinia amylovora suppress plant immunity by binding calcium, thus, preventing MAMP-induced signaling in the plant host (Aslam et al., 2008).
The EPS from P. chlororaphis O6 functions similarly to the EPS from other plant-associated bacteria, e.g., Pantoea agglomerans, Burkholderia gladioli, and P. putida, in the activation of defense mechanisms (Ortmann et al., 2006; Park et al., 2008; Sandhya et al., 2009; Santaella et al., 2008). These findings suggest that it is feasible to develop formulations containing EPS from certain plant-associated bacteria as ŌĆ£green formulationsŌĆØ to boost plant health during crop production. Such formulations would overcome the problem of requiring live microbial cells to induce stress tolerance. Applications of EPS-based formulations maybe especially valuable to aid in alleviating the stress imposed on crops by water shortage caused by higher temperatures and altered precipitation due to climate change.
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






