Occurrence of Bacterial Stem Rot of Ranunculus asiaticus Caused by Pseudomonas marginalis in Korea
Article information

Abstract
In December 2016, stem rot symptoms were observed on Persian buttercup (Ranunculus asiaticus) plants in Chilgok, Gyeongbuk, Korea. In the early stage of the disease, several black spots appeared on the stem of infected plants. As the disease progressed, the infected stem cleaved and wilted. The causal agent was isolated from a lesion and incubated on Reasoner’s 2A (R2A) agar at 25°C. Total genomic DNA was extracted for phylogenetic analysis. Based on the 16S rRNA gene analysis, the isolated strain was found to belong to the genus Pseudomonas. To identify the isolated bacterial strain at the species level, the nucleotide sequences of the gyrase B (gyrB) and RNA polymerase D (rpoD) genes were obtained and compared with the sequences in the GenBank database. As the result, the causal agent of the stem rot disease was identified as Pseudomonas marginalis. To determine the pathogenicity of the isolated bacterial strain, it was inoculated into the stem of healthy R. asiaticus plant, the inoculated plant showed a lesion with the same characteristics as the naturally infected plant. Based on these results, this is the first report of bacterial stem rot on R. asiaticus caused by P. marginalis in Korea.
Introduction
Pseudomonas spp. are naturally widespread in nature and nutritional studies have shown that they can utilize a broad range of organic and inorganic compounds as a sole source of carbon and energy (Stanier et al., 1996). Members of the genus Pseudomonas can survive in any environment with a temperature range of 4-42°C and a pH between 4-8 that contains simple or complex organic compounds (Moore et al., 2006). Pseudomonas spp. have been isolated from varied environments including soil, water, plants, clinical specimens, and marine habitats (Peix et al., 2009; Silby et al., 2011). Numerous Pseudomonas spp. are plant pathogens that can infect various hosts. For example, P. marginalis causes bacterial soft rot on lily bulbs (Hahm et al., 2003), P. syringae pv. syringae causes bacterial spot disease on green pumpkins (Park et al., 2016), and P. viridiflava is a pathogen of Arabidopsis in the Midwestern United States (Jakob et al., 2002).
Ranunculus asiaticus is an important ornamental species that is mainly cultivated in the countries surrounding the Mediterranean Sea but is also cultured in Israel, the Netherlands, the United States, and Japan (Beruto and Debergh, 2004). R. asiaticus is grown outdoors during the fall and winter for spring blooming and its life cycle is well adapted to the Mediterranean climate. It can be propagated by seeds or by tuberous roots (Margherita et al., 1996; Meynet, 1993). Bacterial leaf spot on Ranunculus asiaticus caused by Xanthomonas campestris was observed in California (Azad et al., 1996). In Korea, R. asiaticus was reported to be infected with downy mildew disease caused by Peronospora sp. (Choi et al., 2013) and stem rot caused by Sclerotinia sclerotiorum (Han et al., 2015).
In this article, we describe the identification and characterization of the causal agent of stem rot of R. asiaticus isolated in Korea. A polyphasic examination of the isolate was done by using morphological, biochemical and molecular data, including a phylogenetic analysis based on the 16S rRNA, gyrase B (gyrB), and RNA polymerase D (rpoD) gene sequences. As far as we know, this study is the first molecular and biochemical study of Pseudomonas marginalis associated with stem rot of R. asiaticus in Korea.
Materials and Methods
Bacterial isolation and culture conditions
Tissue sections from lesions on diseased leaves were first disinfected with 70% ethanol, then cut and crushed using a mortar and pestle in sterile distilled water and serially diluted. One hundred microliters of each dilution was spread onto Reasoner’s 2A (R2A) agar plates (Difco, Detroit, MI, USA) and incubated at 25°C for 1 week. On the 103-diluted plate, 30-40 colonies appeared. One white-to-yellowish and transparent colony, designated Ra3, was purified by transferring it onto a fresh plate and incubating it under the same conditions. Strain Ra3 was routinely cultured on R2A agar at 25°C and was maintained as a glycerol suspension (20%, w/v) at -70°C. Growth was assessed on R2A agar (Difco, Detroit, MI, USA), Luria-Bertani agar (LB; Difco), and potato dextrose agar (PDA; Difco) plates, and King’s medium B (KB) plates. The isolate was deposited in the Korean Collection for Type Cultures (KCTC 62486) (Korea Research Institute of Bioscience and Biotechnology, Jeollabuk-do, Korea).
Biochemical characteristics
The Gram stain reaction of the bacterium was determined using the staining (Smibert and Krieg, 1994) and non-staining (Buck, 1982) methods, and its oxidase activity was indicated by the development of a violet-to-purple color following the addition of 1% (w/v) tetramethyl-p-phenylene diamine (BioMérieux, Craponne, France). Fluorescent pigment was tested on King’s medium B (proteose peptone no. 3, 20 g; K2HPO4 · 3H2O, 2.5 g; MgSO4 · 7H2O, 6 g; glycerol, 15 ml; agar, 15 g; and distilled water, 1 l). Enzyme activities, the assimilation of carbon sources, and other physiological characteristics were determined using the API 20NE, API 32 GN, and API 50 CH kits according to the manufacturer’s instructions (BioMérieux).
Genomic DNA extraction, PCR amplification, and sequence analysis
For the phylogenetic analysis, genomic DNA was extracted using the hexadecyl trimethyl ammonium bromide (Sigma- Aldrich, St. Louis, MO, USA) purification method as described previously (Ausubel et al., 1994). The 16S rRNA gene sequence of strain Ra3 was investigated using the method described by Weisburg et al. (1991) with the universal bacterial primers 27F and 1492R. The gyrB and rpoD genes were analyzed as described previously (Yamamoto and Harayama, 1995, 1998; Yamamoto et al., 1999). The primers UP-1E and APrU were used to amplify gyrB, while the primers 70F and 70R were used for rpoD. The partial 16S rRNA, gyrB, and rpoD gene sequences were compiled using the SeqMan software (DNASTAR, Madison, WI, USA).
Phylogenetic analysis
The 16S rRNA, gyrB, and rpoD gene sequences were aligned using the Clustal X computer program (Thompson et al., 1997). Their phylogenetic neighbors were identified and pairwise 16S rRNA gene sequence similarities with closely related species were calculated using the EzBioCloud server (Yoon et al., 2017). Sequence similarities for 16S rRNA, gyrB, and rpoD were analyzed using the BLAST search program at the NCBI website (http://blast.ncbi.nlm.nih.gov/Blast.cgi). Phylogenetic trees were reconstructed not only from the data sets of gyrB and rpoD, but also from the combined nucleotide sequences of these two genes (gyrB and rpoD), assuming that the analysis using a longer sequence would result in a better resolution and reliability. The 17 related taxa were obtained from GenBank and edited using the BioEdit program (Hall, 1999). These strains are listed in Table 1. Multiple sequence alignments were performed using Clustal X program. Gaps and the 5’ and 3’ ends of the alignments were edited manually in BioEdit (Hall, 1999). Evolutionary distance matrices were generated as described by Kimura (1980), and phylogenetic trees were constructed using the neighbor-joining (Saitou and Nei, 1987), maximum-likelihood (Felsenstein, 1981), and maximum-parsimony (Fitch, 1971) algorithms in the MEGA7 program (Kumar et al., 2016), with bootstrap values calculated based on 1,000 replications.

Isolates used in this study and their GenBank accession numbers. The newly generated sequences are indicated in bold
Pathogenicity test.
The pathogenicity was test as descripted previously (Li et al., 2007). Healthy R. asiaticus plants were used for the pathogenicity test. Bacterial colonies of the Ra3 strain were cultured on R2A agar for 48 h at 25°C and suspended in sterile distilled water. The cells were washed, re-suspended in 10 ml sterile distilled water, and counted in a hemocytometer under a light microscope (BX-50, Olympus, Tokyo, Japan), the number of cells per ml was calculated, then diluted into about 1×108 colony-forming units/ml. The healthy R. asiaticus plants were inoculated with 200 μl bacterial suspensions on their stems using a syringe 10 mm below the surface and wounds were made on their stems. Control stems were treated in the same manner but only with sterile water. Each data point represents six replicates. The experiments were repeated three times with similar results. The treated plants were incubated in a moist chamber at a temperature of 25°C for 1 day, then cultured under normal conditions, and the symptoms were checked after 3 days.
Results
Disease symptoms
Stem rot symptoms were observed on R. asiaticus three days after the inoculation with the bacterial isolate (Fig. 1). In the early stage of the disease, several black spots appeared on the stem of the infected plants. As the disease progressed, the infected stem cleaved and wilted. The infect stem lesion location shows water-soaking and brown rot. Rot symptoms developed along the outer stem at early stage and gradually into inner parts of stem. However, no disease symptoms were observed on the stems of the control R. asiaticus.

Symptoms of stem rot on Ranunculus asiaticus. (A) The stem of Ranunculus asiaticus was inoculated by Pseudomonas marginalis Ra3; Left: Stem rot symptoms after inoculation; Right: Healthy stems (control). (B, C) Stem rot disease occurred naturally on the stem of Ranunculus asiaticus. Red arrows indicate the diseased areas.
Isolation and identification of causal agents
The isolated cells were Gram-negative, aerobic, rod-shaped, lacked flagella, and measured 0.8-1.0 μm wide and 1.5-2.3 μm long. Colonies grown on R2A agar at 25°C for 2 days were white-to-yellowish, transparent, circular, convex, smooth, and slimy. Growth occurred on R2A, LB, and PDA agar. Colonies were fluorescent on King’s medium B. Growth was observed at 4°C and 37°C, but not at 41°C. The strain was positive for oxidase and casein hydrolysis. The phenotypic characteristics that identified strain Ra3 as being similar to other P. marginalis strains are listed in Table 2. It showed positive results for the utilization of L-arabinose, D-galactose, gluconate (weakly), glucose, 2-ketogluconate, mannitol, propionate (weakly), sorbitol, sucrose, and trehalose, but negative results for the utilization of cellobiose and L-rhamnose. The biochemical characteristics are almost identical to those of a previously described strain (Kim et al., 2002), as shown in Table 2.

Comparison between the characteristics of P. marginalis Ra3 and those of the P. marginalis isolate reported by Kim et al. (2002)
Phylogenetic analysis
Comparative 16S rRNA gene sequence (1,400 bp) analyses showed that strain Ra3 is phylogenetically affiliated to the members of the genus Pseudomonas calculated using the EzBioCloud (Yoon et al., 2017), with sequence similarities as follows: P. grimontii CFML 97-514T (99.85%), P. rhodesiae CIP 104664T (99.71%), and P. marginalis ATCC 10844T (99.70%). Since it is difficult to identify the closest species in the genus Pseudomonas using the 16S rRNA sequence alone, other gene sequences have been used as markers, such as gyrB, rpoD (Yamamoto et al., 2000), oprI (De Vos et al., 1998), and rpoB (Ait Tayeb et al., 2005). In the present study, to identify the isolated bacterial strain at the species level, the nucleotide sequences of the gyrase B (gyrB) and RNA polymerase D (rpoD) genes (911 bp and 810 bp, respectively) were obtained and compared with the GenBank database. A phylogenetic tree was obtained using the neighbor-joining method with the combined gyrB and rpoD nucleotide sequences of 13 species of Pseudomonas (a total of 17 strains). This analysis was performed under the assumption that using the gyrB and rpoD sequences in combination would give a more accurate estimate of the phylogeny than using either sequence alone. The phylogenetic position of the new isolate revealed that strain Ra3 appeared within the cluster comprising members of the genus Pseudomonas, and was grouped with several P. marginalis strains with a high bootstrap value of 100%, indicating that strain Ra3 is fully consistent with the species P. marginalis (Fig. 2). The 16S rRNA, gyrB, and rpoD gene sequences of strain Ra3 have been deposited under the accession numbers LC375532, LC379203, and LC379204, respectively.
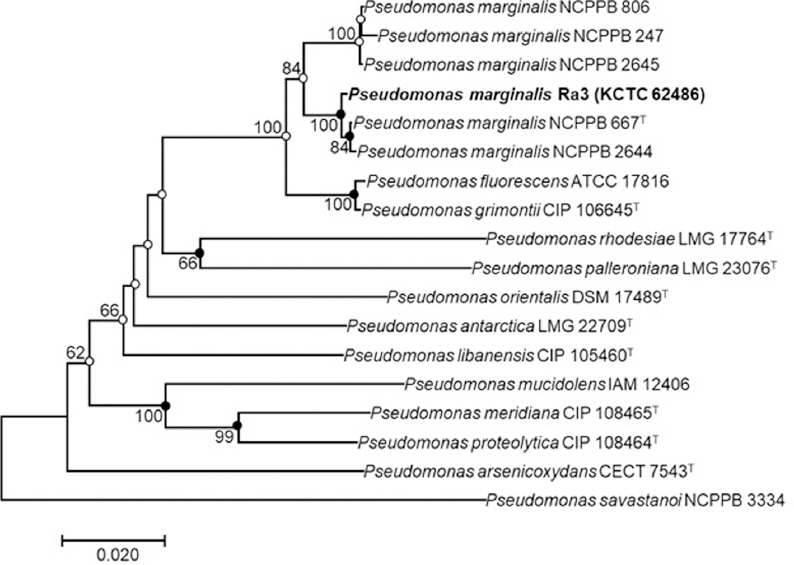
Neighbor-joining phylogenetic tree based on an alignment using the gyrB and rpoD gene sequences in combination. The phylogenic analysis shows the position of Pseudomonas marginalis Ra3 among related strains of Pseudomonas spp. Bootstrap values (based on 1,000 replicates) greater than 50% are shown at branch points. Filled circles indicate that the corresponding nodes were also recovered in trees generated with the maximum-likelihood and maximum-parsimony algorithms. Open circles indicate that the corresponding nodes were also recovered in the tree generated with the maximum-likelihood algorithm. The tree was rooted using Pseudomonas savastanoi NCPPB 3334 as an outgroup. Bar, 0.02 substitutions per nucleotide.
Discussion
The bacterial isolate obtained from stem rot lesions on R. asiaticus was identified as P. marginalis based on morphological, biological, and phylogenetic analyses. The disease symptoms were observed after 3 days of incubation at 25°C. A phylogenetic analysis based on the 16S rRNA gene sequence suggested that Ra3 related to the genus Pseudomonas, and further studies based on the combined gyrB and rpoD gene sequences revealed that strain Ra3 belongs to P. marginalis at the species level. The morphological and biological characteristics of Ra3 were also identical to those of previously described strains of P. marginalis. Phenotypic, morphological, and physiological characteristics as well as the 16S rRNA, gyrB, and rpoD gene sequence analyses indicated that the isolate Ra3 represents a new strain of the species P. marginalis. P. marginalis pv. marginalis is known to cause rot of faba bean (Vassilev, 1998), cauliflower (Obradovic et al., 2002), and cucurbits (Ghobakhloo et al., 2002), leaf spot of philodendron (Schollenberger, 2005), and soft rot of potato (Li et al., 2007) and onion (Achbani et al., 2014). In Japan, Ranunculus production is affected by two diseases, namely mottle caused by a potyvirus (Fujimori et al., 1996) and soft rot caused by P. marginalis pv. marginalis (Kanehira et al., 1997). In Korea, R. asiaticus was reported to be infected with disease of persian buttercup downy mildew caused by Peronospora sp. (Choi et al., 2013) and sclerotinia stem rot caused by S. sclerotiorum (Han et al., 2015). However, there have been no previous reports of P. marginalis infecting R. asiaticus in Korea. Further studies will be needed to develop a molecular method for the rapid detection of P. marginalis, to identify the relative susceptibility of available cultivars, and to study approaches for controlling this disease.
References
Notes
Conflicts of Interest
No potential conflict of interest relevant to this article was reported.