

서론
우리나라에서 배는 수출용 과수로서 중요성이 매우 크다. 국내에서 제일 많이 재배하는 품종은 신고인데, 배 검은별무늬병균 Venturia nashicola에 대해서 감수성이다 보니, 이 품종을 재배하는 농가의 피해가 심각하다(Kwon 등, 2010). 배 검은별무늬병은 4월 하순부터 6월까지 잎과 가지, 열매가 피해를 입기 때문에 기상에 따라 차이는 있지만 해마다 10회에서 18회 이상 약제 방제를 하지 않으면 피해는 더욱 심각하게 발생한다. 현재 등록된 방제 살균제는, 예방살균제, benzimidazole계 살균제, 병원균의 에르고스테롤 생합성을 억제하거나 미토콘드리아의 호흡을 억제하는 살균제 등이 알려져 있다.
이 중 benzimidazole계 살균제는 침투이행성 효과를 갖는 살균제로서, 과수와 작물, 잔디 등에서 Botrytis, Monilinia, Colletotrichum, Sclerotinia, Podosphaera 등이 일으키는 다양한 식물병을 방제하기 위해서 사용한다(Bradley 등, 2006; Chen 등, 2014; Hwang 등, 2010; Sedláková와 Lebeda, 2008). Benzimidazole계 살균제는 식물병원진균의 β-tubulin이라는 단백질에 결합하여 미세소관 형성에 필요한 α, β-dimer의 생성을 저해하기 때문에, 결과적으로 병원균의 세포분열을 억제함으로써 병원균의 생장을 억제하게 된다(Davidse와 Flach, 1977; Martin, 1997). 하지만 작용점이 매우 특이하여 사용한 지 2년 후부터 살균제에 대해서 저항성을 보이는 병원균이 발생하였다(Albertini 등, 1999; Jones와 Walker, 1976; Ross와 Newberry, 1985). 사과 검은별무늬병균인 Venturia inaequalis에 대해서는 1975년에 뉴질랜드에서 benzimidazole계 살균제에 대한 저항성이 보고되었으며, 배에서 검은별무늬병을 일으키는 V. nashicola에서도 1977년에 일본에서 저항성의 발현이 보고되었다(Hartill, 1986; Ishii와 Yamaguchi, 1977). 국내에서도 오이와 인삼에서 분리한 Botrytis cinerea와 포도에서 분리한 Colletotrichum gloeosporioides 등에서 저항성이 보고되었다(Hwang 등, 2010; Kim 등, 2009). Benzimidazole계 살균제 저항성은 병원균의 β-tubulin에서 아미노산 하나가 다른 아미노산으로 치환되어 살균제의 부착이 이루어지지 않기 때문에 나타난다(Ma와 Michailides, 2005). Benzimidazole계 살균제에 대해서 저항성인 여러 종류의 식물병원성 곰팡이의 β-tubulin 유전자를 분석한 결과, 6, 50, 167, 198, 200, 240번의 아미노산 치환이 확인되었다(Albertini 등, 1999; Baraldi 등, 2003; Davidson 등, 2006; Jung 등, 1992; Koenraadt 등, 1992; Ma 등, 2003; McKay 등, 1998; Qiu 등, 2011; Trkulja 등, 2013; Yin과 Xiao, 2013; Zhan과 Huang, 2007). 이처럼 식물병원성 곰팡이에서 β-tubulin 유전자의 일부 아미노산이 치환되면 β-tubulin 단백질에 부착하는 benzimidazole계 살균제의 부착이 억제되어, 살균제를 처리하여도 β-tubulin 단백질에 부착하지 못하고 β-tubulin 단백질이 α-tubulin 단백질과 정상적으로 결합하기 때문에 미세소관을 형성하면서 병원균 생장을 저해하지 못하게 된다. 이처럼 benzimidazole계 살균제는 작용점이 되는 단백질의 아미노산 치환으로 인하여 병원균이 살균제에 대하여 저항성을 나타내게 된다(Yarden과 Katan, 1993). Benzimidazole계 살균제 저항성인 식물병원균에서 저항성과 관련된 β-tubulin 유전자의 치환 부위는 매우 다양한 위치가 보고되어 있으나, 198번째의 아미노산이 glutamic acid에서 alanine으로 치환되는 경우가 가장 많다. 하지만 식물병원균의 종류, 저항성의 정도 등에 따라서 치환되는 위치와 아미노산의 종류가 다르기 때문에 유전자 분석이 필요하다. 사과 검은별무늬병균인 V. inaequalis에서는 198번째 아미노산이 glutamic acid에서 alanine, glycine, lysine 등으로 치환되었으며, 배 검은별무늬병균인 V. pirina에서는 alanine으로 치환되었다(Koenraadt 등, 1992).
하지만 국내 일부 지역에서는 아직도 benzimidazole계 살균제를 사용하고 있는데도 불구하고, 포장에서 저항성 모니터링이 체계적으로 실시된 적은 없다. 따라서 본 논문에서는 한천희석법을 통해서 동양배에 검은별무늬병을 일으키는 V. nashicola의 저항성 검정을 실시하였고, β-tubulin 유전자 분석을 통하여 benzimidazole계 살균제 저항성과 아미노산의 치환 관계를 조사하였다.
재료 및 방법
실험에 사용한 균주와 병원균 분리
Benzimidazole계 살균제에 대한 저항성 검정을 위하여 일본 농업생물유전자은행에서 분양 받아 순천대학교에서 보관하던 V. nashicola 5개의 균주와 2014년 원주, 나주, 옥천 등지의 배 과수원에서 채집한 V. nashicola 30균주를 사용하였다. 채집한 병든 식물체로부터 병원균을 단포자 분리하였다. 병든 식물체 표면에 형성된 분생포자를 수확하여 100 μg/ml의 streptomycin을 첨가한 멸균증류수에 1×104개/ml가 되도록 조절하였다. 병원균의 분생포자 현탁액의 포자밀도를 물한천 배지를 부운 Petri 접시 한 장당 300개의 분생포자가 접종될 수 있도록 밀도를 조절하여 부어주었다. 광학현미경으로 발아한 분생포자를 확인한 후, 한 개의 분생포자를 100 μg/ml의 streptomycin을 첨가한 potato dextrose agar (PDA) 배지에 접종하고 20°C의 암조건에서 10주간 배양하였다. 이렇게 얻은 순수배양체는 4°C에서 보관하며 실험에 사용하였다. 실험에 사용하기 위해서는 보관 균주를 새로운 PDA 배지에 접종하여 동일한 조건에서 10주간 배양한 후 살균제 배지에 접종하는 접종원으로 사용하였다.
실험에 사용한 살균제
Benzimidazole계 살균제인 carbendazim (a.i. 60%, WP), benomyl (a.i. 50%, WP)과 N-phenylcarbamate계인 diethofencarb와 carbendazim의 혼합제(a.i. 25%/25%, WP)를 실험에 사용하였다. 각 살균제는 PDA 배지에서 최종농도가 0.032, 0.16, 0.8, 4.0, 20.0, 100.0 μg/ml가 되도록 첨가하였다.
V. nashicola에 대한 살균제의 균사생장 억제 효과 조사
실험에 사용한 3종류의 살균제를 살균증류수에 현탁한 후, PDA 배지에 정해진 농도가 되도록 맞추어 첨가하였다. 세균의 오염을 방지하기 위해서 300 μg/ml의 streptomycin을 배지에 같이 첨가하였다. 접종을 하기 위하여 준비한 V. nashicola의 균사 선단에서 균사 조각(가로×세로, 1×1 mm)을 떼어 내어 살균제를 첨가한 PDA 배지에 접종하고, 20°C의 암상태에서 4주간 배양하였다. 살균제의 효과는 4주간 배양한 후 살균제 배지에서 자란 병원균의 균총의 직경과 무처리 배지에서 병원균 균총의 직경을 조사하고, 아래 식에 의해서 균사생장 억제효과를 구하여 비교하였다.
병원균의 gDNA 추출
gDNA 추출을 위하여 병원균을 PDA 배지에 접종하여 20°C의 암조건에서 4주간 배양한 후, V. nashicola의 균사를 수확하여 -70°C에서 12시간 동안 동결건조하였다. 멸균한 유리 막대를 사용하여 동결건조한 균사를 고르게 분쇄하고, Kang 등(2005)의 방법으로 gDNA를 추출하였다. 추출한 gDNA는 50 μl의 멸균증류수에 녹인 후 5 mg의 RNase를 처리하여 37°C에서 30분 반응시켜 RNA를 제거하고, 65°C에서 1시간 동안 보관하여 RNase를 불활성화시켰다. 추출된 gDNA는 -20°C의 냉동고에서 보관하며 실험에 사용하였다.
V. nashicola의 β-tubulin 유전자 증폭과 염기서열 분석
Btub14F (5’-AAC CAA ATT GGT GCT GCC TTC TG-3’)와 BtubR (5’-TGG AGG ACA TCT TAA GAC CAC G-3’)을 사용하여 β-tubulin 유전자를 증폭하였다. PCR mixture에는 dNTP 2.5 μl (각 2 mM), 10× Taq buffer 2.5 μl, primer 각각 1 μl (각각 10 pmol), DNA 2 μl (25 ng/μl), Taq polymerase 0.1 μl (5 U/μl) 멸균증류수 15.9 μl가 되도록 하였다. PCR 반응은 1분간 94°C에서 denaturation, 30초간 58°C에서 annealing, 1분간 72°C에서 extention을 35회 반복하였다. 반응을 완료한 산물은 1.5% TAE agarose gel에서 100 V로 40분간 전기영동한 후 UV illuminator에서 band를 확인하였다. PCR로 얻어진 β-tubulin 유전자의 증폭산물은 PCR Expin kit (GeneAll, Seoul, Korea)를 이용하여 정제하였으며, 정제된 PCR 산물을 마크로젠사(Macrogen, Seoul, Korea)에 염기서열 분석을 의뢰하였다.
결과 및 고찰
배 검은별무늬병균의 benzimidazole계 살균제에 대한 반응
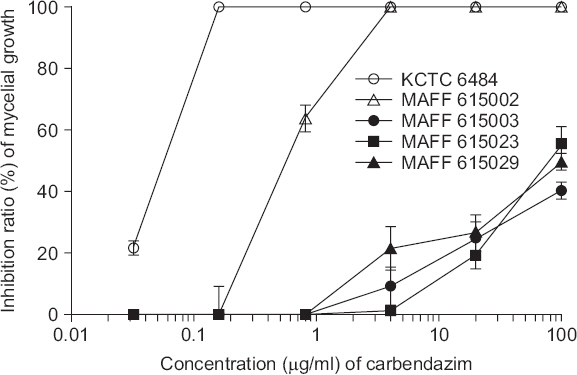
실험에 사용한 V. nashicola 5개 균주의 살균제에 대한 반응은 다양하게 나타났다. Table 1과 Fig. 1에서 보는 것과 같이 감수성인 KCTC6484 균주의 carbendazim에 대한 EC50값은 0.053 μg/ml였으며, carbendazim의 농도가 0.16 μg/ml인 PDA 배지에서는 균사가 전혀 생장하지 못하였다. 하지만 저저항성을 보이는 V. nashicola MAFF615002의 EC50값은 1.154 μg/ml였으며, 4.0 μg/ml의 carbendazim을 첨가한 PDA 배지에서 균사가 전혀 생장하지 못하였다. Carbendazim에 대해서 고저항성인 3개 균주의 EC50값은 모두 100 μg/ml 이상이었으며 가장 높은 농도인 100 μg/ml가 첨가된 배지에서도 병원균의 균사가 생장하였다. Quello 등(2010)은 미국 인디애나주의 사과 검은별무늬병균을 분리하여 한천희석법으로 benzimidazole계 살균제인 benomyl과 thiophanate-methyl에 대한 반응을 조사하여 감수성, 저저항성, 중저항성, 고저항성으로 분류하였다. 감수성은 살균제의 농도가 0.5 μg/ml인 PDA 배지에서 전혀 생장하지 못하는 균주들이고, 0.5 μg/ml 배지에서는 생장하지만 5.0 μg/ml 배지에서 생장하지 못하는 균주들을 저저항성으로 구분하였다. 중저항성은 5.0 μg/ml 배지에서는 생장하지만 50.0 μg/ml 배지에서 생장하지 못하는 균주들이고, 고저항성은 50.0 μg/ml의 배지에서조차 생장이 가능한 균주들이었다. 본 실험에서 사용한 5개 균주 중에서 KCTC6484는 감수성 균주이지만 나머지 4균주는 Quello 등(2010)의 기준에 의하면 MAFF615002는 저저항성 균주이고, MAFF615003을 비롯한 나머지 세 균주는 모두 고저항성 균주로 구분할 수 있었다. 하지만 병원균에 따라서는 benzimidazole계 살균제에 대한 저항성 기준이 다를 수가 있다. Taiwan에서 열대 작물에 탄저병을 일으키는 C. gloeosporioides는 각 균주의 EC50값을 가지고서 benomyl에 대한 저항성 기준을 나누면서 감수성은 EC50값이 10 μg/ml 이하, 중저항성은 10-100 μg/ml 사이, 그리고 중저항성은 100-500 μg/ml 사이, 고저항성은 500 μg/ml 이상으로 구분하였다(Chung 등, 2010). 미국 캘리포니아에서는 갈색 부패 증상을 보이는 핵과류에서 39개의 Monilinia fructicola 균주를 분리하여 benomyl에 대한 감수성과 저항성을 구분한 결과, 감수성의 EC50값은 0.03-0.05 μg/ml 사이, 저저항성은 0.61-0.96 μg/ml 사이, 그리고 고저항성을 50 μg/ml 이상으로 구분하였다. M. furucticola는 동일한 benzimidazole 그룹의 살균제라고 할지라도 살균제에 따라서 그 반응이 다르게 나타났는데, thiphanate-methyl에 대해서는 감수성의 EC50값의 범위는 0.26-0.74 μg/ml였으며, 저저항성은 2.38-6.42 μg/ml 사이였고, 고저항성은 benomyl과 동일하게 50 μg/ml 이상이었다(Ma 등, 2003). 사탕무 점무늬병을 일으키는 Cercospora beticola는 EC50값이 1.0 μg/ml 이하인 균주를 감수성으로, 1.0-100 μg/ml 사이의 균주를 저저항성으로, 100-1,000 μg/ml 사이를 중저항성, 그리고 1,000 μg/ml 이상인 균주를 고저항성으로 구분하였다(Trkulja 등, 2013). 이처럼 살균제에 대한 감수성과 저항성의 구분은 병원균에 따라서, 또 동일한 기작을 갖는 그룹에 속하는 살균제라고 하더라도 살균제의 종류에 따라서 다르기 때문에 정확한 저항성의 기준이 필요하다. 실험한 5개의 균주를 가지고 benzimidazole계 살균제에 대한 저항성 기준을 설정하였다. 실험한 균주는 감수성, 저저항성, 고저항성 등 3개의 그룹으로 나눌 수 있었는데, 0.16 μg/ml의 carbendazim 배지에서 전혀 생장하지 못하는 균주를 감수성으로 분류하였다. 저저항성은 4.0 μg/ml carbendazim 배지에서 전혀 생장하지 못하며, 고저항성은 100 μg/ml carbendazim 배지에서 생장이 가능한 균주이다. Benzimidazole계 살균제에 속하는 benomyl에 대해서도 5균주는 동일한 반응을 보였는데, V. nashicola KCTC6484는 감수성, V. nashicola MAFF615002는 저저항성, 그리고 나머지 3균주는 모두 고저항성을 보여 carbendazim에 대한 반응과 동일함을 알 수 있었다. 결국 배 검은별무늬병균은 동일한 benzimidazole계 살균제에 속하는 carbendazim과 benomyl에 대해서 교차 저항성 관계가 있음을 알 수 있다.
Table 1
EC50 value* of Venturia nashicola causing pear scab to benzimidazole fungicides and the mixture of carbendazim and diethofencarb
| Isolates† | Carbendazim | Benomyl | Carbendazim+diethofencarb |
|---|---|---|---|
| V. nashicola KCTC 6484 | 0.053 | 0.009 | 0.022 |
| V. nashicola MAFF 615002 | 1.154 | 0.813 | 0.595 |
| V. nashicola MAFF 615003 | 59.123 | >100 | 0.327 |
| V. nashicola MAFF 615029 | >100 | >100 | 0.150 |
| V. nashicola MAFF 615023 | >100 | >100 | >100 |
* EC50 value (μg/ml) means the effective concentration reducing the mycelial growth of each isolate of V. nashicola by 50%.
† Each isolate of V. nashicola was inoculated on a potato dextrose agar (PDA) medium amended with or without fungicide at indicated concentration, and incubated at 20°C under the dark condition for 4 weeks. Colony diameter was measured one month later. Inhibitory ratio (%) of mycelial growth was calculated as follows;
Fig. 1
Controlling efficacy of carbendazim against the mycelial growth of Venturia nashicola isolates. All isolates used in this experiment were incubated on potato dextrose agar medium amended with or without carbendazim by indicated concentrations at 20°C for 4 weeks. After that, the diameter of colony was measured to investigate the inhibitory ratio of the fungicide.
원주, 나주, 옥천에서 분리한 30균주의 carbendazim에 대한 반응을 살펴보면 Fig. 2에서 보는 것과 같이 모든 균주가 100 μg/ml carbendazim 배지에서 생장하여, 국내에서 분리한 모든 균주는 carbendazim에 대해서 고저항성이었다.
Fig. 2
Effect of carbendazim on the mycelial growth of Venturia nashicola on potato dextrose agar (PDA) amended with the fungicide. Isolates of V. nashicola were collected in Wonju (A), Naju (B), and Okcheon (C). Isolates of V. nashicola were inoculated on PDA with carbendazim by an indicated concentration and PDA without the fungicide as an untreated control. All isolates were incubated at 20°C for 4 weeks, and the diameter of them was measured. The inhibitory efficacy (%) of the fungicide against the mycelial growth was calculated by comparing the colony diameter of V. nashicola on PDA with carbendazim to that on PDA without the fungicide.

Carbendazim/Diethofencarb 혼합제의 균사생장 억제효과
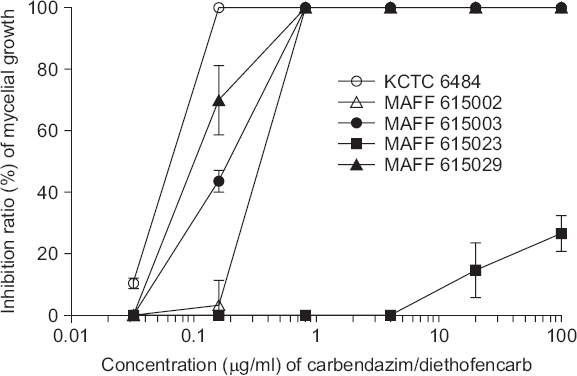
실험에 사용했던 5개 균주 중에서 carbendazim에 대해서 감수성 균주인 KCTC6484와 저저항성 균주인 MAFF615002, 그리고 고저항성 균주 중에서 MAFF615023을 제외한 MAFF615003과 MAFF615029 두 균주는 모두 carbendazim과 diethofencarb의 혼합제에 대해서 감수성이었다(Fig. 3). 감수성인 V. nashicola KCTC6484의 합제에 대한 EC50값은 0.022 μg/ml로 가장 낮은 EC50값을 보여주었으며, 0.16 μg/ml의 혼합제가 첨가된 PDA 배지에서 균사가 전혀 생장하지 못한 반면에, carbendazim에 대해서 저저항성인 MAFF615002와 고저항성인 MAFF615003과 MAFF615029 등 3개 균주의 EC50값은 각각 0.595, 0.327, 0.150 μg/ml였으며, 혼합제가 0.8 μg/ml 첨가된 PDA 배지에서 전혀 생장하지 못하였다. 하지만 고저항성이었던 MAFF615023은 100 μg/ml의 PDA 배지에서조차 생장하였으며, EC50값도 100 μg/ml 이상으로 carbendazim뿐만 아니라 carbendazim과 diethofencarb 혼합제에 대해서도 저항성인 것으로 나타났다.
Fig. 3
Effect of the fungicide mixing carbendazim and diethofencarb on the mycelial growth of Venturia nashicola. Inhibitory efficacy of the fungicide against the mycalial growth of V. nashicola isolates was investigated on potato dextrose agar (PDA) amended with or without the fungicide by an indicated concentration through an agar dilution method. All isolates of V. nashicola inoculated on PDA were incubated at 20°C for 4 weeks. Colony diameters were measured after 4 weeks of incubation. Inhibition ratio of the mycelial growth was defined as the diameter of a mycelial colony on PDA with the fungicide by an indicated concentration per colony diameter on unamended PDA×100.
혼합제의 구성성분인 carbendazim과 diethofencarb는 서로 역상관 교차저항성 관계에 있는 살균제로서 carbendazim에 대해서 저항성인 균주는 diethofencarb에 대해서 감수성이며, 감수성인 균주는 저항성을 보인다(Elad 등, 1988). 따라서 포장에서 식물병원균이 benzimidazole계 살균제에 대해서 저항성이 발현되어, 저항성균과 감수성균의 집단이 혼재해 있을 경우 이 혼합제를 처리하면 carbendazim 저항성 병원균은 diethofencarb에 의해서 생장이 억제되고, 감수성인 균주는 carbendazim에 의해서 억제되기 때문에 저항성이 발현된 병원균이 집단을 이루고 있는 포장이라고 해도 방제가 가능하다(Ishii와 Takeda, 1989). 하지만 토마토, 포도, 오이 등에서 분리한 잿빛곰팡이병균 중에서 역상관 교차저항성 관계에 있는 carbendazim과 diethofencarb 모두에 대해서 저항성인 균주가 발견되었다(Pappas, 1997). Koenraadt 등(1992)도 사과와 서양배에서 검은별무늬병을 일으키는 V. inaequalis와 V. pirina를 분리하여 benomyl에 대한 저항성 정도를 저저항성(low resistance), 중저항성(medium resistance), 고저항성(high resistance), 극저항성(very high resistance) 등으로 구분하고, 이 각각의 그룹이 benomyl과 N-phenylcarbamate계에 속하는 methyl N-(3,5-dichlorophenyl) carbamate (MDPC)와 역상관 교차저항성 관계가 있는지를 조사하였다. V. inaequalis의 경우 저저항성 그룹의 균주들을 제외한 다른 그룹의 저항성 균주들은 MDPC에 대해서 감수성 반응을 보였으며, V. pirina의 경우에는 고저항성과 극저항성 그룹에 속하는 균주만이 MDPC에 대해서 역상관 교차저항성을 보여주었다. 본 실험에서는 Koenraadt 등(1992)과 다르게 diethofencarb와 carbendazim의 혼합제를 사용하여 실험하였다. 따라서 carbendazim에 대해서 감수성인 균주와 저항성인 균주 모두가 합제에 대해서는 감수성을 보여야 한다. 5균주 중에서 carbendazim에 대해서 감수성인 2균주와 저항성인 2균주 모두가 역상관 교차저항성 관계에 있는 carbendazim과 diethofencarb의 합제에 대해서는 감수성을 보였는 데 비해서, V. nashicola MAFF615023은 carbendazim과 합제 모두에 대해서 저항성이었다. Ishii와 Takeda (1989)도 V. nashicola 균주 중에서 carbendazim과 역상관 교차저항성 관계에 있는 화합물인 N-phenylformamidoxime계 N-(3,5-dichloro-4-propynyloyphenyl)-N’-methoxyformamidine (DCPF)에 대해서 모두 저항성을 보이는 균주가 있으며, 방사선 동위원소인 14C가 라벨되어 있는 carbendazim과 DCPF를 사용하여 부착 실험을 한 결과, 두 살균제 모두에 대해서 저항성인 균주에 잘 부착하지 않음을 보고하였다. 하지만 국내 3지역의 배 과수원에서 채집한 V. nashicola 균주들은 Fig. 4에서 보는 것과 같이 4.0 μg/ml의 혼합제를 첨가한 배지에서 전혀 생장하지 못하여서 국내에서는 아직 혼합제에 대한 저항성균의 발현은 없는 것으로 생각하였다.
Fig. 4
Effect of the carbendazim (A) and the mixture (B) of carbendazim (Car) and diethofencarb (Die) on the mycelial growth of Venturia nashicola isolated from orchards located on Wonju of Kangwon, Naju of Jennam, and Okcheon of Chungbuk. Inhibition ratio of the mycelial growth was defined as the diameter of a mycelial colony on potato dextrose agar (PDA) with the fungicide by an indicated concentration per colony diameter on unamended PDA×100. Colony diameters were measured 4 weeks after incubation at 20°C.

β-Tubulin 유전자의 염기서열 분석
Btub14F/BtubR primer를 사용하여 실험에 사용한 5균주로부터 1,264 bp의 β-tubulin 유전자를 증폭하였으며, 증폭한 유전자의 염기서열을 분석한 결과, Fig. 5에서처럼 carbendazim에 대한 저항성 여부에 따라서 변화하였다. 감수성인 V. nashicola KCTC6484와 저저항성인 MAFF615002는 198번째의 아미노산 코돈이 GAG로 glutamic acid였는데, carbendazim에 대해서 고저항성인 MAFF615003과 MAFF615029는 GCG로서 alanine이었으며, carbendazim과 diethofencarb의 혼합제에 대해서 저항성인 MAFF615023은 AAG로 lysine인 것으로 밝혀졌다. 포장에서 채집한 균주 또는 실험실에서 인위적으로 돌연변이를 일으켜 선발한 benzimidazole계 살균제 저항성 균주들의 β-tubulin 유전자를 분석한 결과 다양한 위치에서 아미노산 치환이 결국 살균제 저항성을 유도하였다(Ma와 Michailides, 2005). 이처럼 β-tubulin에서 아미노산이 치환되면 살균제가 β-tubulin에 부착되지 못하기 때문에 감수성이 저하하고 저항성이 발현된다(Ishii와 Takeda, 1989). 그런데 아미노산이 치환되는 위치와 치환되는 아미노산의 종류에 따라서 저항성 정도에도 차이가 있다. Benzimidazole계 살균제에 대해서 고저항성인 Monolinia fructicola는 β-tubulin의 198번째 아미노산이 glutamic acid에서 alanine으로 치환되었지만, 저저항성인 M. fructicola는 6번째 아미노산이 histidine에서 tylosine으로 치환되었다(Ma 등, 2003). 동일한 저항성이지만 V. inaequalis의 경우는 M. fructicola와 다르게 고저항성 균주의 198번째 아미노산이 glutamic acid에서 lysine으로 치환되었다(Koenraadt 등, 1992). 본 실험에서 사용하였던 V. nashicola 5균주는 198번째의 아미노산이 glutamic acid나 lysine으로 치환되면서 carbendazim에 대해서 저항성을 보였다. 하지만 MAFF615023과 같이 lysine으로 치환된 균주는 benzimidazole계 살균제인 carbendazim뿐만 아니라 역상관 교차 저항성 관계에 있는 N-phenylcarbamate계 diethofencarb와의 혼합제에 대해서도 저항성을 보임으로써 경우에 따라서는 역상관 교차 저항성과 관계없이 저항성이 발현됨을 보여주었다.
Fig. 5
Nucleotide (A) and deduced amino acid (B) sequence of β-tubulin gene from 5 isolates of Venturia nashicola. The mutation site at codon 198 is indicated with a box.

원주, 나주, 옥천 등 국내 세 지역에서 채집한 30개의 V. nashicola 균주 모두는 carbendazim과 carbendazim과 diethofencarb 혼합제에 대해서는 감수성이었다(Table 2). 이들의 β-tubulin을 분석하면 198번째의 아미노산이 다른 보고들과 동일하게 glutamic aicd에서 alanine으로 치환되어 있었지만, 옥천에서 분리한 5개 균주, OC2, OC5, OC8, OC55, OC57만이 alanine이 아닌 GGG인 glycine으로 치환되어 있었다. Glycine 치환은 carbendazim에 대해서는 저항성을 나타내었지만, 혼합제에 대해서는 다른 균주들보다 더 감수성을 보였다. Fig. 4에서 보는 것과 같이 glycine 치환 균주는 0.16 μg/ml의 혼합제 처리에서도 균사가 전혀 생장하지 못하였으며, EC50값 역시 0.03 μg/ml 이하로 나타나 매우 감수성인 균주임을 알 수 있었다(Table 2). 하지만 MAFF615023과 같이 역상관 교차 저항성 관계에 있는 carbendazim과 diethofencarb 혼합제에 대한 저항성을 발견되지 않았으며, 오히려 β-tubulin의 아미노산 치환에 의해서 더 감수성을 보이는 균주가 발견되었다. 이처럼 198번째의 아미노산이 glycine으로 치환되어 carbendazim에 대해서 저항성이 되면서, 역상관 교차 저항성 관계가 있는 carbendazim과 diethofencarb 혼합제에 대해서 감수성이 커지는 경우는 처음 보고되는 결과이다.
Table 2
EC50 value* of Venturia nashicola isolates collected from Wonju, Naju and Okcheon in 2014, and amino acid at 198th of β-tubulin
| Isolates | EC50 value (μg/ml) | 198th codon | Amino acid | |
|---|---|---|---|---|
|
|
||||
| Carbendazim | MixtureCar+Die† | |||
| WJ6 | 12.14 | 0.11 | GCG | Alanine |
| WJ14 | 16.97 | 0.10 | GCG | Alanine |
| WJ15 | >100.00 | 0.37 | GCG | Alanine |
| WJ23 | 43.39 | 0.09 | GCG | Alanine |
| WJ27 | 11.75 | 0.12 | GCG | Alanine |
| WJ32 | 97.76 | 0.43 | GCG | Alanine |
| WJ33 | >100.00 | 0.33 | GCG | Alanine |
| WJ34 | >100.00 | 0.33 | GCG | Alanine |
| WJ37 | 32.37 | 0.36 | GCG | Alanine |
| WJ38 | 14.48 | 0.13 | GCG | Alanine |
| NJ4 | 64.11 | 0.15 | GCG | Alanine |
| NJ5 | 75.10 | 0.06 | GCG | Alanine |
| NJ6 | 61.90 | 0.13 | GCG | Alanine |
| NJ8 | 45.45 | 0.13 | GCG | Alanine |
| NJ9 | >100.00 | 0.40 | GCG | Alanine |
| NJ15 | >100.00 | 0.39 | GCG | Alanine |
| NJ17 | 73.40 | 0.52 | GCG | Alanine |
| NJ20 | 74.13 | 0.10 | GCG | Alanine |
| NJ31 | 39.08 | 0.40 | GCG | Alanine |
| NJ34 | 71.25 | 0.48 | GCG | Alanine |
| OC2 | 43.37 | 0.02 | GGG | Glycine |
| OC5 | 29.37 | 0.02 | GGG | Glycine |
| OC8 | 26.32 | 0.03 | GGG | Glycine |
| OC16 | 39.17 | 0.39 | GCG | Alanine |
| OC30 | 55.05 | 0.34 | GCG | Alanine |
| OC36 | 27.52 | 0.36 | GCG | Alanine |
| OC48 | 19.60 | 0.32 | GCG | Alanine |
| OC54 | 21.58 | 0.33 | GCG | Alanine |
| OC55 | 10.27 | 0.01 | GGG | Glycine |
| OC57 | 19.80 | 0.02 | GGG | Glycine |
Benzimidazole계 살균제는 1970년대부터 사용되기 시작한 침투이행성 살균제로서 적용 범위가 넓어서 많은 식물병의 방제에 사용되어 왔지만, 포장에서 식물병 방제를 위해서 살균제를 처리하면서부터 2년 사이에 저항성균이 출현하기 시작하였다. 이제는 다양한 식물병원균에서 benzimidazole계 살균제에 대한 저항성이 문제가 되고 있다. 국내에서는 배 검은별무늬병의 방제를 위해서 benzimidazole계 살균제가 사용되었지만, 저항성 검정이 이루어지지는 않았다. 하지만 지금도 지역에 따라서는 검은별무늬병 방제 체제 안에서 benzimidazole계 살균제를 처리하고 있기 때문에 저항성 발현에 대한 정확한 검정이 필요한 시기였다. 2014년 국내 세 지역에서 채집한 균주는 30균주 모두가 benzimidazole계 살균제에 대해서 저항성으로 나타나 국내의 검은별무늬병균의 저항성화가 심각한 수준임을 짐작하게 한다. 따라서 전국적인 범위에서 저항성 검정이 이루어져야 하며 포장에서 살균제 저항성 관리가 적극적으로 이루어져야 할 것으로 생각한다.
요약
Benzimidazole계 살균제 저항성을 모니터링하면서, 저항성과 β-tubulin 유전자의 198번째 코돈의 돌연변이와의 관계를 조사하였다. Venturia nashicola의 benzimidazole계 살균제에 대한 반응은 0.16 μg/ml carbendazim 배지에서 전혀 생장하지 못하는 감수성과 4.0 μg/ml 배지에서 생장이 안 되는 저저항성, 100 μg/ml에서조차 생장이 가능한 고저항성으로 구분할 수 있었다. 국내 3개 지역의 과원에서 채집한 30균주의 V. nashicola는 carbendazim에 대해서 모두 고저항성이었다. V. nashicola의 β-tubulin 유전자의 염기서열을 분석한 결과 저저항성 균주에서는 감수성 균주와 염기서열에 차이가 없었지만, 고저항성 균주는 GAG인 glutamic acid가 GCG인 alanine으로 치환되어 있었다. 옥천의 과원에서 분리한 10균주 중에서 5균주는 GGG인 glycine으로 치환되어 있었다. 특히 glycine으로 치환될 경우 carbendazim에 대해서 저항성이 되면서 carbendazim과 diethofencarb 혼합제에 대해서는 감수성이 더 커졌다. 결국 β-tubulin의 198번째 코돈의 돌연변이 E198A와 E198G는 V. nashicola가 benzimidazole계 살균제에 대해서 저항성이 되는 중요한 기작임을 알 수 있었다.
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






