

서론
뿌리혹선충(Meloidogyne spp.)은 식물 뿌리에 절대기생하며 토마토, 고구마, 참외, 오이 등 주요 농작물에 심각한 피해를 야기하는 토양 병원균 중의 하나이다. 전 세계적으로 작물 피해의 약 11%가 선충에 의한 것으로 알려져 있으며(McCart, 2009), 국내의 경우에도 경북 성주, 충남 공주 등의 포장에서 뿌리혹선충에 의한 병 발생 피해사례가 보고되고 있다(Park 등, 2011, 2012). 현재까지 동정된 약 90여 종 이상의 다양한 뿌리혹선충 중에서도 M. incognita, M. arenaria, M. javanica, M. hapla 등은 넓은 기주 범위를 가지며 작물 수확량을 감소시키기 때문에 경제적으로 중요한 뿌리혹선충으로 간주하고 있다(Kim 등, 2011, 2014; Trudgill과 Blok, 2001). 뿌리혹선충에 의해 감염된 식물체의 뿌리에는 거대한 혹이 형성되며, 이로 인해 식물체는 물과 영양분의 흡수나 미네랄의 이동 등에 방해를 받게 되어 정상적인 생장이 어려워진다(Williamson과 Hussey, 1996). 이처럼 혹이 형성된 식물뿌리의 표면, 또는 그 혹의 내부에는 알이 존재하게 되며, 적절한 조건이 만족되면 알에서 J2 단계의 유충이 부화된다(Perry와 Moens, 2006). J2 유충 단계의 뿌리혹선충은 식물체의 뿌리 끝으로 침투하여 세포 사이를 돌아다니다가 유관속주로 침입하여 뿌리혹을 재형성하게 된다(Caillaud 등, 2008).
식물기생선충에 의한 피해를 감소시키기 위해 윤작, 객토, 태양열 소독, 담수처리 등과 같은 물리적 방제를 비롯하여 저항성 식물 품종 재배, 화학적 방제법 등이 고려된다(Park 등, 2012; Perry와 Moens, 2006). 뿌리혹선충병의 화학적 방제 수단으로는 메틸 브로마이드(methyl bromide) 등의 훈증제와 포스티아제이트(fosthiazate) 등의 살선충제가 주로 이용되어 왔다. 하지만 이러한 화학적 방제는 인축에 대한 위험성과 잔류독성, 환경오염, 생태계 교란 등과 같은 다양한 문제점을 노출하고 있으며, 이로 인해 보다 안전한 살선충제의 개발이 요구되고 있다. 환경 친화적으로 식물기생선충을 방제하기 위하여 식물 추출물이나 미생물, 또는 미생물의 이차대사산물을 이용한 방법들이 고려되고 있으며, 동시에 많은 연구들이 진행되어 오고 있다(Akhtar와 Malik, 2000; Kerry, 2000; Oka 등, 2000; Rodriguez-Kabana 등, 1987; Sharon 등, 2001). Pseudomonas fluorescens의 배양여액은 in vitro상에서 선충의 알 부화율 감소 및 M. javanica의 유충에 대한 살선충 효과를 보였으며, 이는 P. fluorescens의 이차대사산물인 2,4-diacetylphloroglucinol에 의한 효과로 밝혀졌다(Siddiqui와 Shaukat, 2003). 또한 Trichoderma harzianum BI과 27종의 식물체로부터 얻은 다양한 essential oil도 M. javanica의 치사율에 영향을 준다는 연구 결과가 있다(Oka 등, 2000; Sahebani와 Hadavi, 2008). 최근 Jang 등(2016)은 Aspergillus niger F22의 배양여액이 M. incognita J2 유충의 치사율 증가 및 선충 알의 부화율 감소 결과를 초래한다고 보고하였으며, 이는 A. niger F22 균주가 생산하는 oxalic acid로부터 기인한 효과라고 보고하였다.
토양에 서식하는 선충은 다른 균류들에 의해 자극 받을 수 있어 토양미생물을 활용한 생물학적 선충방제법이 많은 관심을 받아오고 있다. 방선균은 토양에 서식하는 세균으로, 균사의 형태로 증식하면서 다양한 이차대사산물을 생성한다(Siddiqui와 Mahmood, 1999). 현재까지 알려진 천연 항생물질의 60%-70%가 방선균으로부터 발견되었으며, 1970년대에 그 수가 최대치를 찍은 이후로도 매년 이들이 내는 항생물질이 꾸준히 보고되고 있다(Bérdy, 2005; Watve 등, 2001). Avermectin, bafilomycin, fervenulin, 6’-methyl-fungichromin 등이 방선균에 의해 생산되는 살선충 물질로 알려져 있으며, 이들 물질들은 살선충 활성 이외에도 병원성 곰팡이나 박테리아에 대한 항균 활성을 갖는 것으로 보고되어 있다(Lacey 등, 1995; Putter 등, 1981; Ruanpanun 등, 2011; Zeng 등, 2013). 하지만 이러한 다양한 연구결과에도 불구하고 길항 방선균을 직접 이용한 미생물 농약 개발은 미비한 실정이다.
따라서 본 연구에서는 토양에서 분리한 670여 개의 방선균을 대상으로 뿌리혹선충(M. incognita)에 대한 살선충 활성 조사를 실시하였고, 이 중 가장 우수한 활성을 갖는 Streptomyces flavogriseus KRA15-528 균주를 선발하였다. 선발된 균주에 대한 in vivo와 in vitro 살선충 활성 조사를 통해 친환경 살선충제로서의 개발 가능성을 검토하였다.
재료 및 방법
방선균의 배양 및 배지조건
2015년도에 다양한 지역의 토양으로부터 방선균을 분리하였으며, 분리된 방선균의 살선충 활성을 조사하고자 다음과 같은 배지 및 배양조건을 이용하였다. 방선균은 Bennett’s 한천배지(1% glucose, 0.1% yeast extract, 0.2% Bacto-peptone, 0.1% beef extract, 2% agar) (Hesseltine 등, 1954)에 도말하여 고체 배양한 후, 형성된 단일 콜로니를 tryptic soy broth 배지에 접종하고 28°C, 150 rpm에서 2일 동안 전배양하였다. 전배양한 방선균은 glucose-soybean meal-starch (GSS) 배지(1% soluble starch, 2% glucose, 2.5% soybean meal, 0.1% beef extract, 0.4% yeast extract, 0.2% NaCl, 0.025% K2HPO4, 0.1% CaCO3) (Kim 등, 1989)에 1% (v/v)가 되도록 접종하고 28°C, 150 rpm에서 7일 동안 진탕 배양하여 살선충 활성 조사에 사용하였다.
뿌리혹선충의 증식 및 분리
Hwang 등(2014)에 의해 분리, 동정된 뿌리혹선충은 온실(25°C-5°C)에서 ‘서광 ’ 품종의 토마토에 증식시켜 실험에 사용하였다. 원예용 상토(High; Punong, Gyeongju, Korea)를 직경 5 cm의 플라스틱 컵에 넣고 토마토를 파종하였다. 3주간 재배한 유묘를 플라스틱 포트에 이식한 뒤 뿌리혹선충의 알을 토마토 한 주당 5,000개씩 접종하였으며, 6주 동안 재배하여 혹이 심하게 형성된 토마토 뿌리를 채취하였다.
뿌리에 생긴 혹과 난낭으로부터 뿌리혹선충 유충(J2)을 분리하기 위해 혹이 형성된 토마토 뿌리를 물로 세척한 후 1 cm 크기로 절단하였다. 토마토 뿌리가 0.5% 차아 염소산나트륨(sodium hypochlorite) 용액에 잠길 정도로 넣은 후, 분쇄기로 60초간 분쇄하였다. 분쇄한 시료는 230 mesh 체를 이용하여 찌꺼기를 거르고 550 mesh 체를 이용하여 알을 수집하였다(Barker 등, 1985). 체에 모인 선충 알은 멸균수로 3번 세척한 뒤 부화 억제 활성 조사에 이용되거나, Baermann funnel 방법을 이용하여 선충 알로부터 유충을 분리하였다(Southey, 1986).
방선균 배양여액이 뿌리혹선충 유충과 알에 미치는 영향
GSS 배지에서 7일 동안 배양한 방선균 배양액을 4°C, 6,500g 조건에서 10분 동안 원심분리하여 얻어진 상등액을 사용하여 유충에 대한 살선충 활성과 알 부화 억제 활성 조사를 하였다. 먼저, 유충에 대한 살선충 활성을 조사하기 위해 선충 현탁액을 96-well tissue culture plate (Becton Dickinson, Franklin Lakes, NJ, USA)의 각 well에 넣은 다음 방선균 배양여액의 농도가 10%, 5%, 2.5%, 1.25%가 되도록 처리하고 well당 최종 부피가 100 μl (약 50마리 J2)가 되도록 하였다. 시료가 균일하게 섞이도록 30초 동안 교반하였으며, 선충의 산소결핍과 건조를 방지하기 위하여 상대습도 100%의 플라스틱 통에 넣은 후 25°C에서 배양하였다. 실험은 3반복을 수행하였으며, 시료처리 48시간, 72시간, 96시간 후 광학현미경(CKX41; Olympus, Tokyo, Japan)하에서 관찰하여 직선으로 뻗어 움직임이 없는 선충을 죽은 선충으로, 움직임이 있는 선충을 살아있는 선충으로 판단하여 살선충률을 조사하였다(Abbott, 1925). 알 부화 억제 활성을 조사하기 위해 유충에 대한 살선충 활성을 조사하기 위한 방법과 동일한 방법을 사용하였다. 선충 알을 각 well에 약 50개씩 넣은 다음 방선균 배양여액의 농도가 10%, 5%, 2.5%로 되도록 처리하고 well당 최종 부피가 100 μl가 되도록 하였다. 배양조건은 유충에 대한 살선충 활성 실험에 사용했던 조건과 같으며, 선충 알 부화율은 시료처리 후 3일, 7일, 9일째 조사하였다.
살선충율(%)=[(처리구의 치사율대조구의- 치사율)/(100대조구의-치사율)]×100
알 부화율(%)=[유충의 수/(알의 수+유충의 수)]×100
분리 균주의 형태학적 및 분자계통학적 분석
선발된 KRA15-528 균주의 형태학적 특성은 Bennett ’s 한천배지에 접종하여 28°C 배양기에서 10일 동안 배양한 뒤 관찰하였다. 주사전자현미경을 통하여 균주의 포자형태와 크기를 관찰하기 위해 멸균된 금속을 Bennett ’s 한전배지에 비스듬히 끼워 넣은 뒤 금속과 배지가 맞닿는 선을 따라 KRA15-528 균주를 도말하였다. 28°C에서 10일간 배양하여 금속을 타고 올라온 균체를 주사전자현미경(MIRA3; TESCAN, Seoul, Korea)을 통해 관찰하였다(Williams와 Davies, 1967).
분리 균주의 16S rRNA 유전자 염기서열을 바탕으로 분자계통학적 분석을 실시하였다. KRA15-528을 Bennett’s 액체배지 5 ml에 접종하여 28°C, 150 rpm 조건으로 3일간 배양하였다. 배양액을 4°C, 6,500g 조건으로 10분간 원심분리하여 얻은 균체로부터 cetyltrimethylammonium bromide를 이용하여 genomic DNA를 추출하였다(Tripathi와 Rawal, 1998). 16S rRNA 부분의 증폭을 위하여 universal primer set (27F, 5’-AGAGTTTGATCMTGGCTCAG-3’; 1492R, 5’-TACGGYTACCTTGTTACGACTT-3’)를 이용하였으며, 중합 효소 연쇄반응은 Ex Taq polymerase (Takara, Ohtsu, Japan)를 사용하여 94°C 5분 1회, 94°C 30초, 58°C 30초, 72°C 30초에서 34회, 72°C에서 7분간 1회 반응시켰다. 확인된 밴드는 GeneAll Expin™ PCR purification kit (GeneAll, Seoul, Korea)의 방법에 따라 정제한 후 염기서열 분석을 위해 시퀀싱 분석 서비스(Macrogen, Daejeon, Korea)를 이용하였다. 분석된 염기서열은 BLASTn (http://www.ncbi.nlm.nih.gov) 프로그램을 통해 비교 분석되었다. 분자 계통학적 계통수의 작성은 MEGA 프로그램 패키지를 사용하여 수행하였으며(Tamura 등, 2013), 분석을 위한 염기서열 정렬은 CLUSTAL W (Larkin 등, 2007)를 이용하였다. 분자계통도는 거리행렬법 중의 하나인 neighbor-joining 방법(Saitou와 Nei, 1987)을 사용하여 구성하였고, bootstrap 분석은 1,000회 수행하였다.
배양여액 용매 분획들의 살선충 활성
방선균 배양여액의 용매 분획에 따른 살선충 활성을 조사하기 위해 배양여액과 동량의 에틸 아세테이트와 부탄올을 이용하여 순차적으로 각각 2회씩 용매 분획하여 감압 농축하였다. 에틸 아세테이트, 부탄올, 물로부터 추출물 0.01, 0.45, 0.59 g씩을 확보하였으며, 각 추출물을 100 mg/ml 농도가 되도록 메탄올(에틸 아세테이트와 부탄올 추출물) 또는 50% 메탄올(물 추출물)에 용해하였다. 96-well tissue culture plate의 각 well에 뿌리혹선충 2령 유충(약 50마리/99 μl)을 치상한 후 각각의 시료를 처리하여 최종 농도가 250, 500, 1,000 µg/ml가 되도록 하였다. 무처리 대조구로는 1% 메탄올을 사용하였으며, 실험은 3회 반복 수행되었다. 시료 처리 후 48시간 뒤, 살선충률을 조사하였다.
In vivo 뿌리혹선충병 방제 활성 검정
오이를 기주로 이용하여 KRA15-528 균주 배양여액의 뿌리혹선충병 방제활성을 조사하였다. 온실(25°C-5°C)에서 5일 동안 키운 ‘백미다다기’ 품종의 오이를 멸균한 모래 100 g이 담긴 직경 7 cm의 플라스틱 포트에 이식하였다. 토마토에서 증식된 뿌리혹선충 알을 수거하여 각 포트당 5,000알을 접종하였다. 접종 후 한 시간 뒤, KRA15-528 균주의 배양여액을 오이 뿌리 주변에 10 ml씩 관주처리하였으며, 일주일 후 같은 방식으로 배양여액을 한번 더 토양관주 처리하였다. 대조구로는 선충탄(a.i. 5% fosthiazate) 2,000배 희석액을 1회 처리하였으며, 무처리구로는 멸균수를 이용하였다. 실험은 각 처리구당 5개의 식물체를 사용하여 2반복을 수행하였고, 선충 알 접종 2주 후에 뿌리혹 지수를 조사하였다. 뿌리혹 지수는 오이뿌리에 형성된 혹의 정도에 따라 0-4의 지수를 사용하여 5단계로 조사하였다(0, 0%; 1, 1%-25%; 2, 26%-50%; 3, 51%-75%; 4, 76%-100%) (Taylor와 Sasser, 1978).
결과 및 고찰
방선균의 in vitro 살선충 활성과 알 부화 억제 활성
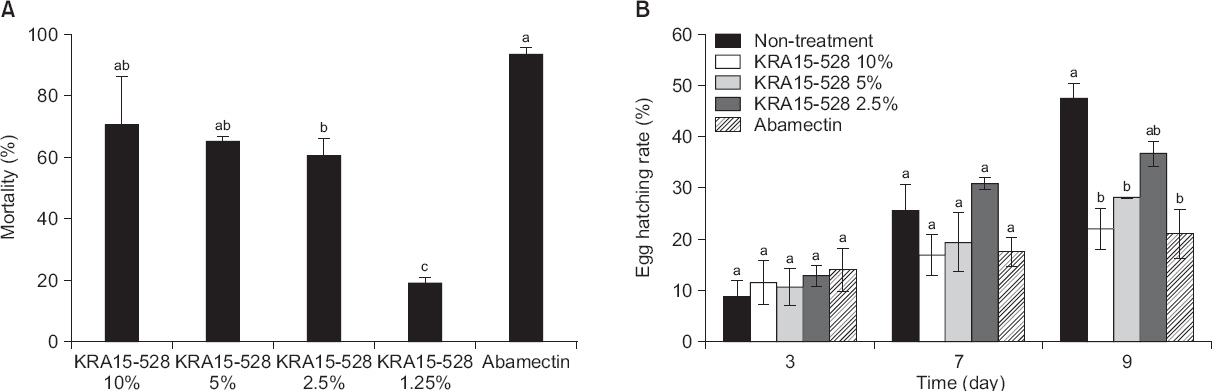
살선충 활성을 가지는 균주의 선발을 위하여 670여 개의 방선균 배양여액을 이용하여 96-well tissue culture plate에서 스크리닝을 수행하였다. 약 670개의 방선균 중 KRA15-528이 가장 높은 살선충 활성을 가지는 균주로 선발되었다. KRA15-528 균주를 선충 현탁액에 처리 후, 48시간째에 관찰하였을 때 10% 처리구에서는 71%, 5%에서는 65%, 2.5%에서는 61%의 높은 살선충률을 비롯하여 농도 의존적인 결과를 보여주었다. 또한 1.25% 처리구에서도 19%의 활성을 나타냈다(Fig. 1A). 배양여액 처리 후, 72시간 및 96시간째에서도 48시간째 관찰한 살선충률과 비슷한 살선충률을 나타내어 지속적인 살선충 활성을 보임을 알 수 있었다(data not shown). Jang 등(2015)은 S. netropsis AN10065 균주의 배양액 추출물을 선충에 5% 농도로 처리 시 72시간 후 20% 이하의 살선충률이 나타났다고 보고하였으며, Kim 등(2011)은 S. sampsonii KK1024 배양액을 10% 농도로 처리했을 때 21%의 살선충률을 보고하고 있다. 하지만 본 연구에서는 KRA15-528 균주의 배양여액 10% 처리 시 71%의 높은 살선충 활성을 확인하였다(Fig. 1A). 배양조건 등 실험적 환경의 차이를 감안한다고 하더라도 앞선 연구 결과와 비교하여 본 실험에 사용된 Streptomyces sp. KRA15-528 균주가 in vitro에서 우수한 살선충률을 보여주고 있음을 알 수 있었다.
Fig. 1
Effects of culture filtrates of Streptomyces flavogriseus KRA15-528 on J2 mortality (A) and egg hatching (B) of Meloidogyne incognita. Values are mean±standard deviation of three replicates. Means with the same letter are not significantly different (P<0.05) according to Duncan’s multiple range test. Abamectin (μg/μl) was used as a control. (A) The number of dead nematodes were counted 48 hours after treatment. Mortality rates (M) were corrected using Abbott’s formula: M=[(Mt-Mc)/(100-Mc)]×100, where Mt means mortality percentage in treatment and Mc means mortality percentage in control. (B) The number of hatched eggs were counted 3, 7, and 9 days after treatment. Egg hatch rate (EH) was calculated as follows: EH=[J/(E+J)]×100, where J and E mean the juveniles and eggs of M. incognita, respectively.
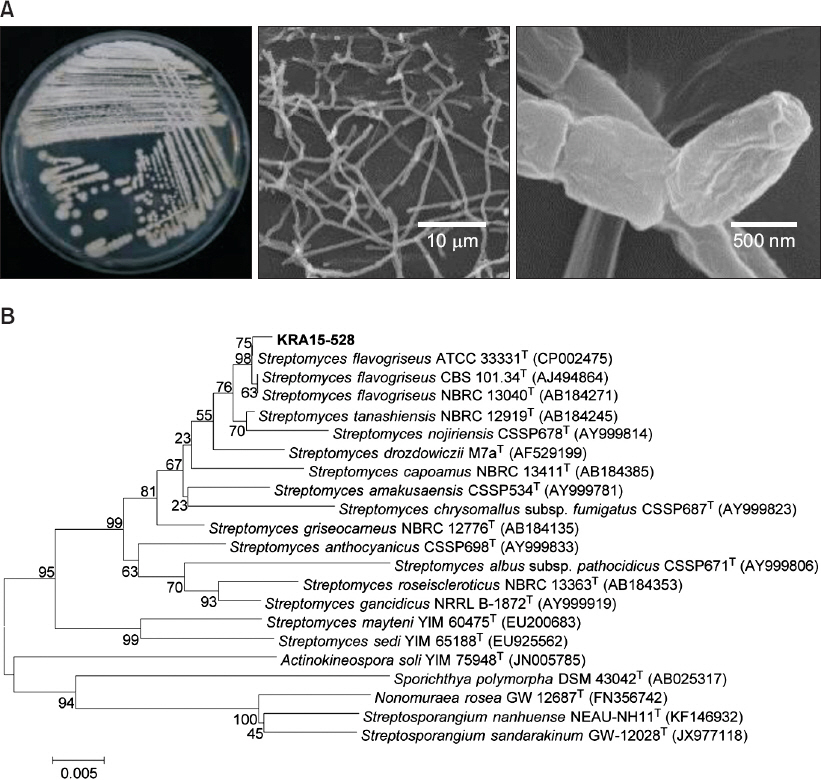
KRA15-528 균주의 형태학적 특성 및 계통수 분석
KRA15-528 균주는 Bennett ’s 한천배지에 도말하였을 때 사상형의 균사는 회색 및 노란색을 띠며, 주사전자현미경으로 관찰 시 균사의 끝부분에 분절된 형태의 원통 모양 포자가 관찰되었다(Fig. 2A). 포자의 크기는 약 0.7-1 μm였다(Fig. 2A). 16S rRNA 유전자 염기서열을 이용한 분자 계통학적 분석 결과, S. flavogriseus ATCC 33331과 99.86%, S. tanashiensis NBRC 12919와 98.49%, S. nojiriensis CSSP678과는 96.83%의 염기서열 유사도를 보였다(Fig. 2B). 이에 상기에서 분리된 균주를 S. flavogriseus KR15-528로 명명하였다.
Fig. 2
Morphological feature and phylogenetic analysis of Streptomyces flavogriseus KRA15-528. (A) Left picture shows a culture plate of KRA15-528 grown on Bennett’s agar medium 28°C for 10 days. Middle and right pictures are scanning electron micrograph images of hyphae and spore surface of KRA15-528, respectively. (B) Phylogenetic analysis was performed based on 16S rRNA gene sequence of KRA15-528 and 21 species of the genus Streptomyces. Neighbor-joining method was used for this analysis, and numbers at nodes indicate levels of bootstrap support (%) by 1,000 resampled datasets. National Center for Biotechnology Information accession numbers of each sequence are in parentheses. Bar, 5 substitution per 1,000 nt.
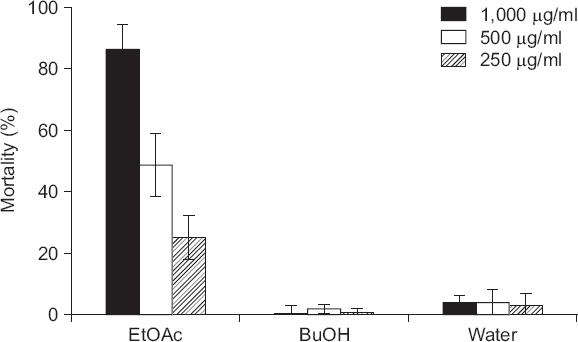
용매 분획별 살선충 활성
KRA15-528 균주 배양여액으로부터 살선충 활성 물질의 특성을 파악하고자 용매 분획된 에틸 아세테이트, 부탄올, 물층을 각각 선충 현탁액에 처리해 보았다. 처리 후 48시간에서 관찰한 결과, 에틸 아세테이트 추출물 처리구에서 높은 살선충 활성이 나타났다(Fig. 3). 부탄올 추출물과 물 층은 1,000 µg/ml 농도로 처리 시 살선충률이 각각 4%와 8%로 매우 낮았다(Fig. 3). 에틸 아세테이트 추출물의 경우 250, 500, 1,000 µg/ml 처리구에서 각각 30%, 53%, 91%의 농도 의존적인 살선충률을 보여주었다(Fig. 3). 또한, 72시간 및 96시간 후에도 지속적인 살선충 효과를 보여주었다(data not shown). 이들 결과를 바탕으로, 에틸 아세테이트 추출물에는 KRA15-528 균주가 생성하는 살선충 활성 물질이 포함되어 있으리라 생각되며, 향후 살선충 활성 물질의 동정에 대한 연구가 필요하다.
Fig. 3
Nematicidal activity of two organic layers and aqueous layer from culture filtrate of Streptomyces flavogriseus KRA15-528. The number of dead nematodes were counted 48 hours after treatment. Mortality rate (M) was calculated as follows: M=[(Mt-Mc)/(100-Mc)]×100, where Mt means mortality percentage in treatment and Mc means mortality percentage in control. Values are mean±standard deviation of three replicates. Abamectin (μg/μl) was used as a control.
현재 대표적인 살선충제로 판매되고 있는 abamectin은 avermectins B1a와 B1b의 혼합제이며, 이들 물질들은 S. avermitilis의 이차대사산물로 알려져 있다(Lasota와 Dybas, 1991; Roberts와 Hutson, 1999). 또한, KRA15-528 균주와 동일 종인 S. flavogriseus는 protease, polycyclic xanthone계의 항생물질인 SIIA-C2191A와 SIIA-C2191B 등의 이차대사산물을 생산한다는 보고가 있으며, S. flavogriseus ACTK2 균주의 부탄올 추출물은 Staphylococcus aureus, Bacillus subtilis, Enterococcus aerogenes, Fusarium proliferatum 등에 대하여 항균활성을 가진다는 연구결과가 보고되어 있다(Dezfully와 Ramanayaka, 2015; Ghorbel 등, 2014; Zhao 등, 2003). 본 연구에서 선발된 S. flavogriseus KRA15-528 균주는 벼 잎집무늬마름병, 토마토 잿빛곰팡이병, 토마토 역병, 밀 붉은녹병, 고추 탄저병 등에 방제활성이 없는 것으로 관찰되었지만(data not shown), 항균활성에 대한 방선균의 넓은 스펙트럼을 고려한다면 다양한 식물병원균을 이용한 방제활성의 조사가 필요하리라 생각된다.
방선균은 다양한 생리활성물질을 생산한다. 이러한 물질은 크게 세포내 물질과 세포외 물질로 나눌 수 있다. 현재까지 보고된 방선균에 의해 생산되는 살선충 물질들, 예를 들면, avermectins, bafilomycin, lecanicidins 등은 세포내 물질로 알려져 있다(Burg 등, 1979; Lacey 등, 1995; Nonaka 등, 2000). 본 연구에서 S. flavogriseus KRA15-528 균주의 배양여액으로부터 확인된 살선충 활성은 활성물질이 세포외 물질일 것으로 추정되며, 동시에 상기 균주로부터 살선충 활성을 갖는 세포내 물질을 동정한다면 고활성을 보이는 미생물 살선충제 개발이 가능해지리라 생각된다.
균주 배양여액의 뿌리혹선충에 대한 in vivo 실험
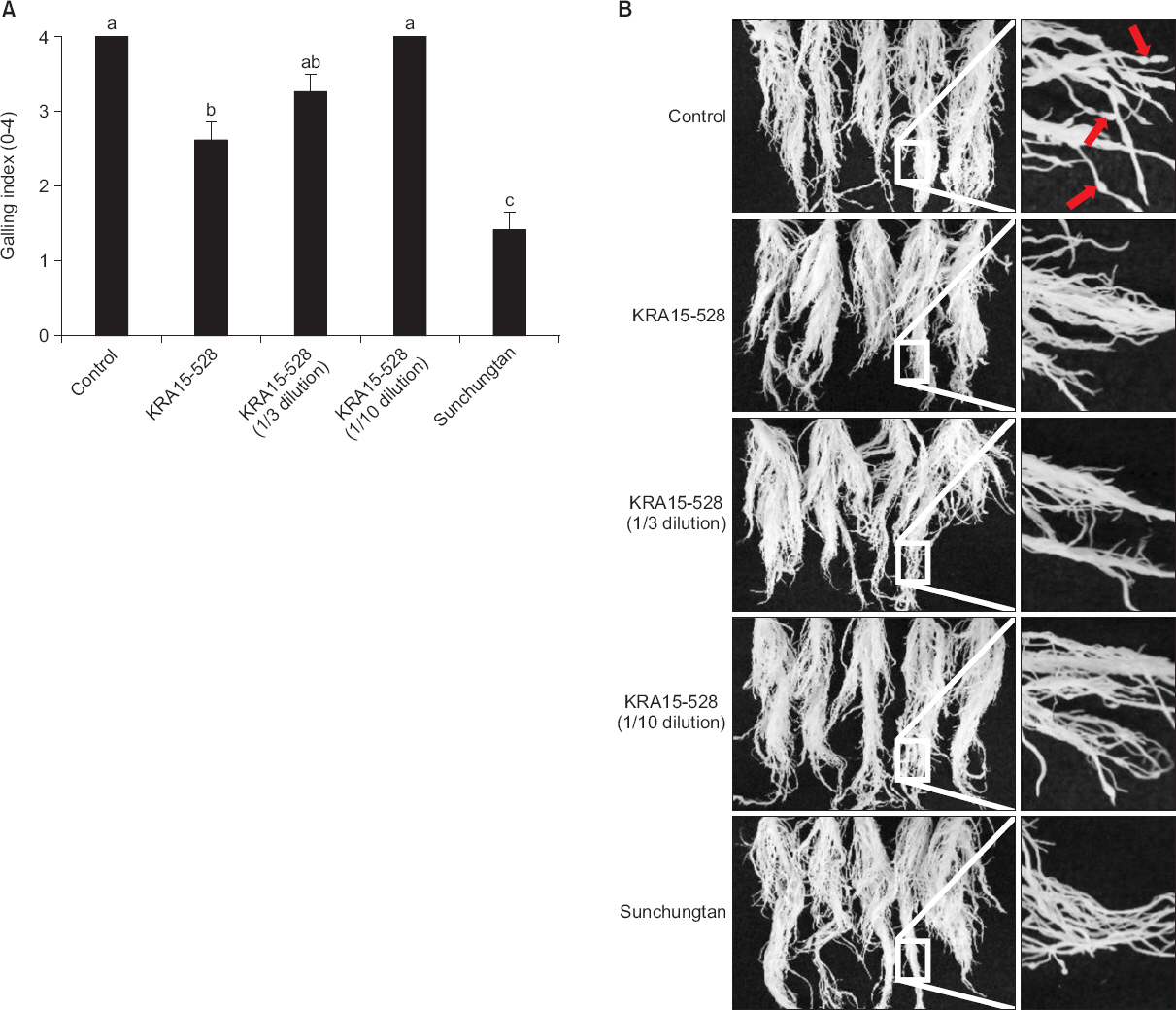
KRA15-528 균주가 뿌리혹 선충에 대해 식물병방제 활성을 가지는지 알아보기 위하여 오이에 선충을 처리한 후 KRA15-528 균주의 배양여액을 처리하였다. 배양여액의 원액, 1/3배, 1/10배 희석액 처리구에서 각각 2.6, 3.3, 4.0의 뿌리혹형성지수를 보였으며, 무처리구에서는 4.0의 뿌리혹형성 지수를 나타내었다. KRA15-528 배양여액(원액)은 무처리구와 유의적으로 차이를 보였으나, 1/3배와 1/10배 희석액은 유의적인 차이가 없었다(Fig. 4A) (Taylor와 Sasser, 1978). 또한, 오이 이외에도 토마토를 대상으로 한 실험결과에서도 KRA15-528 균주 배양여액에 의한 방제활성이 확인되었으며, 모든 in vivo 실험상의 처리구에서 약해는 발견되지 않았다(data not shown).
Fig. 4
Inhibition activity of Streptomyces flavogriseus KRA15-528 to gall formation on cucumber roots. (A) At two weeks after infection of root-knot nematodes, the galling index was assessed according to a 0-4 galling scale; 0=0%, 1=1%-25%, 2=26%-50%, 3=51%-75%, and 4=76%-100% (Taylor and Sasser, 1978). Each value represents the mean±standard deviation of five replications. The experiment was performed twice with similar results (data from a representative experiment are shown). Means with the same letter are not significantly different (P<0.05) according to Duncan’s multiple range test. (B) Galling formation was suppressed by treatment of culture filtrate of KRA15-528. Photographs were taken 2 weeks after inoculation of Meloidogyne incognita. Left panel is magnified pictures of right panel, and red arrows indicate galls formed at roots.
본 연구에서는 670여 개의 방선균 가운데 가장 살선충 활성이 우수한 S. flavogriseus KRA15-528 균주를 선발하였고, 본 균주의 살선충 활성 물질은 에틸 아세테이트로 추출이 가능한 세포외 물질로 확인되었다. 또한 오이를 이용한 온실 시험에서도 뿌리혹선충에 대하여 방제 활성을 보였다. 향후, 본 균주를 이용하여 미생물 살선충제를 개발하기 위해서는 살선충 활성 물질 분리 및 구조 동정, 최적 발효 공정, 최적의 살선충 활성을 위한 제제 개발 등의 추가적인 연구가 필요하다.
요약
뿌리혹선충에 의한 병 발생은 작물수확량의 감소를 초래하여 중요한 문제로 인식되고 있다. 화학농약의 사용은 선충방제를 위해 효과적인 방법 중의 하나로 고려되지만 환경오염이나 인축에 대한 독성과 같은 문제를 야기하고 있다. 본 연구에서는 친환경 살선충제 개발에 사용될 수 있는 미생물을 선발하고자, 토양으로부터 분리한 670여 개의 방선균을 대상으로 뿌리혹선충에 대한 살선충 활성을 조사하였다. 이들 중 가장 우수한 살선충 활성을 나타내는 KRA15-528 균주를 선발하였으며, 16S rRNA 유전자 염기서열에 기초하여 S. flavogriseus로 동정하였다. S. flavogriseus KRA15-28 균주 배양여액을 최종농도 10%가 되도록 J2 단계의 유충과 선충 알에 각각 처리 시, 48시간 후 71%의 선충 치사율과 9일째에 54%의 선충 알 부화 억제율을 확인하였다. 배양여액을 에틸 아세테이트, 부탄올을 이용하여 용매 분획하여 에틸 아세테이트와 부탄올, 물 추출물에 대한 살선충 활성을 조사한 결과, 에틸 아세테이트 추출물로부터만 높은 살선충 활성을 확인하였다. 1,000, 500, 250 µg/ml의 농도에서 각각 91%, 53%, 30%의 치사율을 나타내었다. 이외에도, 오이에 대한 뿌리혹선충병 방제 활성을 조사한 결과, 배양여액 처리구에서 눈에 띄게 뿌리혹형성이 억제되는 현상을 관찰하였다. 따라서, S. flavogriseus KRA15-528은 뿌리혹선충병을 방제에 사용할 수 있는 미생물제제로서의 가능성이 있음을 확인할 수 있었다.
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






