

서론
콩(Glycine max)은 장미목(Rosales) 콩과(Leguminosae)에 속하는 한해살이 쌍떡잎식물로 한반도를 포함한 아시아 동북부 지역이 원산지로 알려져 있으며, 전 세계적으로 550속 13,000 종, 우리나라에서는 36속 92종이 자생하고 있다. 콩은 단백질과 지방 공급원으로 예로부터 중요시 되는 작물이며, 현재는 공업용 소재로도 각광받고 있는 작물이다(Ann, 2012).
콩의 중요 바이러스인 콩모자이크바이러스(Soybean mosaic virus, SMV)는 Potyviridae 과, Potyvirus 속에 속하는 바이러스로 진딧물을 통해 충매전염 되며, 콩에서 최대 43%의 종자전염이 이루어지는 것으로 보고되어 있다. 이 바이러스는 콩 생산량과 직접적으로 관련 있는 꼬투리 수의 감소, 꼬투리 당 종자수의 감소, 종자의 크기 및 중량의 감소와 콩 껍질의 얼룩생성 등을 초래한다(Domier 등, 2007, 2011). SMV는 국내에 11개 계통들이 보고되어있으며, 저항성 유전자를 극복하는 계통들이 계속적으로 나타나고 있다(Seo 등, 2009).
최근 주로 사용되는 바이러스 진단방법은 Enzyme-Linked Immuno Sorbent Assay(ELISA) 방법과 Reverse Transcription- Polymerase Chain Reaction(RT-PCR) 방법이다. ELISA 방법은 많은 양의 시료를 쉽게 검사 할 수 있지만 검출한계의 문제와 검사결과 판독의 오류 가능성이 존재한다(Lee 등, 2010). RT-PCR 진단방법은 진단 시료수가 많아지면 진단비용과 소요시간, 기술 숙련도 및 별도의 장비들이 많이 필요하다는 단점이 있다(Li 와 Mock, 2005). 최근에는 기존의 유전자 진단법의 단점을 극복하기 위하여 간단하고 빠르게 식물 바이러스를 진단하는 연구가 이루어져왔다. Notomi 등(2000)은 strand displacement activity가 높은 Bst polymerase를 이용하여 등온에서 특정 유전자를 loop 구조로 증폭시키는 Loop-mediated isothermal amplification(LAMP)법을 고안하였다. 일반 PCR의 경우 2개의 primer들을 사용하는 것과는 달리, LAMP 방법은 6개 영역을 인식하는 4개의 primer들을 사용하게 된다. 증폭 효율이 높고, template를 1시간 정도에 109-1010배까지 증폭시킬 수 있으며, 매우 높은 특이성을 나타내므로 목적 DNA 배열의 존재를 증폭산물의 유무로 판정할 수 있다. 즉 기존의 PCR 방법과 유사 하지만 기존의 중합효소 연쇄반응이 변성(denaturation), 접 합(annealing), 신장(extention) 세 가지 단계를 거치면서 온도의 변화를 주어야 하는 반면, 일정한 온도에서 접합 및 신장이 가능하다는 특징이 있다(Blomstrom 등, 2008; Nagamine 등, 2002). RNA 바이러스를 검출하기 위해서는 역전사 과정이 수반 되기 때문에 일반 DNA 바이러스를 검출하는 것보다 더 많은 시간이 소모된다. 하지만, RT-LAMP는 LAMP와 마찬가지로 등온에서 RT과정과 LAMP 반응이 동시에 수행함으로서 진단 시간을 현저히 줄일 수 있는 장점이 있어 이미 다른 분야에서는 RTLAMP가 적용된 바 있다(Fukuta 등, 2003; Parida 등, 2004). 또한 일정한 온도만 유지가 된다면 반응이 이루어지기 때문에 등온 유지가 가능한 항온 수조나 heating block 등 저가의 장비에서도 반응이 가능하여 고가의 Thermocycler 장비가 필요하지 않다. 그리고 SYBR Green I을 사용하여 전기영동없이 육안확인이 가능하기 때문에 유해한 ethidium bromide(EtBr)를 사용하지 않고, 현장 및 기타 어느 장소에서도 병원체에 대한 진단이 가능한 장점을 가지고 있다.
본 연구에서는 특이성, 정밀성, 간편성을 갖춘 분자기법 중 하나인 RT-LAMP 방법을 이용하여 콩의 중요 바이러스인 SMV의 감염 유무를 신속하게 확인할 수 있는 진단법을 개발하고자 하였다.
재료 및 방법
시험재료
본 연구에서 사용된 SMV는 2013년도 전국 주요 콩 재배지에서 채집된 시료와 보유 중인 9개의 SMV 계통을 반응에 이용하였다. 채집된 시료와 분양받은 시료는 초저온냉동 고(-70°C)에 보관하면서 실험에 이용하였다.
전체 RNA 분리
식물체 100 mg을 Lysis 용액(iNtRON bio., Korea)을 이용하여 제공자의 방법에 따라 분리하였다. 전체 RNA는 최종적으로 30 µl의 elution buffer에 녹여 RT-LAMP와 RT-PCR에 이용하였다.
RT-LAMP primer 제작
종 특이적 primer 선발을 위하여 NCBI GeneBank에 등록된 SMV의 11개 계통 G1(Accession no. FJ640977), G2(S42280), G3(FJ640978), G4(FJ640979), G5(AY294044), G6(FJ640980), G7(AY216010), G5H(FJ376388), G6H(FJ640981), G7A(FJ640982), G7H(AY294045)와 미국에서 분리된 SMV 계통 TNP(HQ845735)의 유전정보 염기서열 을 DNASTAR(DNASTAR, Inc., USA) 소프트웨어를 이용하여 비교 분석하였다. 가장 변이가 적은 외피단백질 염기서열 정보를 이용하여 LAMP primer designing software인 Primer Explorer(http://primerexplorer.jp/e/index.html)를 이용하여 RT-LAMP용 primer들을 제작하였다. 4가지로 이루어진 SMV RT-LAMP용 primer들에서 SML-FIP와 SML-BIP는 loop를 형성하는 염기서열 부분 TTTT spacer를 포함하여 각각 42 nt로 설계되었으며, SML-F3와 SML-B3는 inner primer들의 바깥쪽에 위치 하도록 각각 18 nt, 19 nt로 설계하였다(Table 1). 설계된 primer 들은 Bioneer(Daejeon, Korea)에 주문 의뢰하여 합성하였으며, inner primer들에 대해서는 PAGE 정제된 것을 사용하였다.
Table 1
Specially designed set of four target-specific primers (F3, B3, FIP and BIP)
최적 조건 확립
RT-LAMP 반응은 RT-LAMP kit(Eiken Chemicals Co., Japan)와 반응액을 조제하는 방식으로 반응을 실시하였다. RT-LAMP kit의 사용은 제조사의 방법에 따라 실시하였다. 반응액 조성은 바이러스에 감염된 시료의 total RNA 1 µl와 40 pM inner primers, 5 pM outer primers, 8 U Bst DNA polymerase large fragment(New England Biolabs, USA), 1×ThermoPol Reaction Buffer [20 mM Tris-HCl, 10 mM KCl, 10 mM (NH4)2SO4, 2 mM MgSO4, 0.1% Triton X-100, New England Biolabs, USA], 10 mM dNTP, 5% DMSO, DW로 총 20 µl의 반응액을 조성 하였다. 최적 반응 온도와 시간을 확인하기 위하여 온도는 48- 66°C, 시간은 10-120분으로 변화를 주어 반응을 실시하였다. 반응시간 이후에는 80°C에서 10분간 유지시킨 후 반응을 종료 하였다. 각 LAMP 반응이 끝나면 증폭산물들은 2% agarose gel에서 전기영동(HOEFER, USA)하여 EtBr로 염색하여 확인하였다.
Primer의 특이성 확인
primer의 종 특이성 확인을 위하여 콩에서 발생하는 바이러스들에 대해 반응 여부를 확인하였다. 종 특이성에 이용된 6종의 바이러스는 SMV를 포함하여 Soybean yellow mottle mosaic virus(SYMMV), Soybean yellow common mosaic virus(SYCMV), Peanut stunt virus(PSV), Bean common mosaic virus(BCMV)와 Peanut mottle virus(PeMoV)이 다. 또한 보유 중인 SMV 9계통 G1, G2, G3, G5, G5H, G6, G6H, G7과 G7H에 대해서 그 특이성을 확인하였다. 또한, RT-LAMP에 이용되는 네 개의 특이 primer들에 대한 요구 특이성을 확인하기 위하여 네 개의 primer들 중 하나 혹은 두 개 이상을 제외시켜 조성하고 정상적인 증폭산물과 비교하여 반응 여부를 확인 하였다.
민감도 확인
RT-LAMP는 total RNA를 주형으로 사용하는 것으로 전체 RNA내에서 특정 mRNA의 농도를 확인하기 위하여, SMV의 RT-LAMP로 증폭되는 부위를 포함하는 DNA clone을 주형으로 하여 민감도를 확인하였다. DNA clone 합성에 이용된 primer는 SM_LN2(5’-GTA CAA TGC AGT TAA AGA TGA ATA- 3’)과 SM_LC2 (5’-TCT CAA ATT CCT CAG TAG TCC ATA-3’)이다. 이 DNA clone을 주형으로 1×100부터 1×10-7까지 단계적으로 희석하여 RT-LAMP와 RT-PCR을 수행하여 민감도를 비교 확인 하였다.
결과 및 고찰
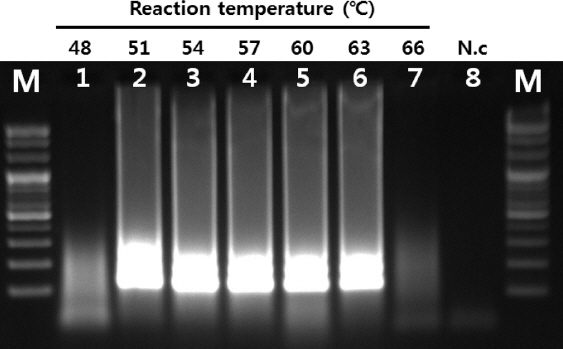
RT-LAMP의 최적 온도를 확인하기 위하여 반응온도를 48-66°C로 각기 등온 조건을 유지하면서 실시하였다. 51°C부터 63°C의 범위에서 증폭이 가능한 것으로 확인되었으며, 최적의 반응을 보인 온도는 57-60°C로 확인되었다(Fig. 1). 이후에 진행된 모든 반응은 58°C 등온 조건에서 실시되었다.
Fig. 1
Optimal reaction temperature of RT-LAMP. LAMP was performed under range of 48 to 66oC. Lane M, 100 bp DNA ladder (Solgent, Korea); lanes 1-7 (48, 51, 54, 57, 60, 63 and 66oC, respectively); lane 8, negative control.
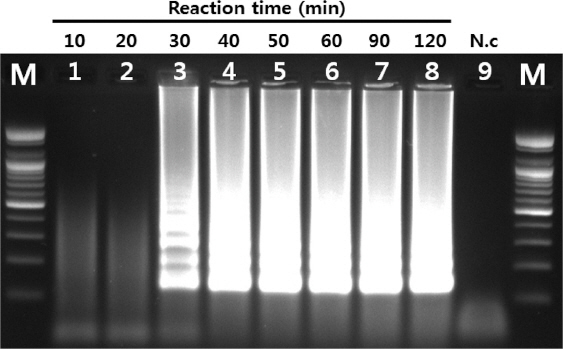
최소 반응시간을 확인하기 위하여 반응시간을 10, 20, 30, 40, 50, 60, 90, 120분으로 하여 증폭 여부를 확인하였다. 최소 30분 이상의 반응시간이 주어질 때 반응이 확인되었으며, 60분 이상이면 최적의 반응을 나타내는 것으로 확인되었다(Fig. 2). 이후에 진행된 모든 반응 시간은 60분으로 실시되었다.
Fig. 2
Optimal reaction time of RT-LAMP. LAMP was performed under range of 10 to 120 min. Lane M, 100 bp DNA ladder; lanes 1-8 (10, 20, 30, 40, 50, 60, 90 and 120 min, respectively); lane 9, negative control.
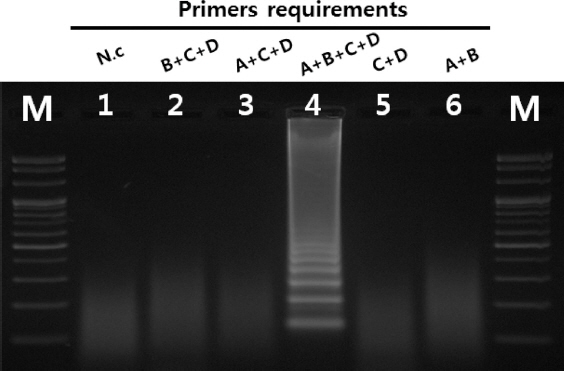
Primer들의 종 특이성 확인을 위하여 콩에서 발생하는 주요 바이러스 6종에 대하여 반응 여부를 확인한 결과 SMV에서만 반응이 확인되어 SMV의 RT-LAMP 진단에 특이적인 primer들로 확인되었다(Fig. 3). 또한 보유 중인 SMV의 9계통 G1, G2, G3, G5, G5H, G6, G6H, G7와 G7H에 대해서 그 특이성이 확인되었다(Fig. 4). 이번 실험에서 이용된 primer들의 경우 SMV의 외피단백질을 기초로 하여 설계되었다. SMV의 외피단백질 유전자의 경우 계통 간 변이가 크지 않으며, 검정한 9계통 모두에서 그 반응의 특이성이 확인되어 모든 계통에 적용하여도 가능할 것으로 예상된다. 4가지로 이루어진 SMV의 RT-LAMP용 primer 들에 대한 요구 특이성을 확인하기 위하여 primer를 하나 또는 두 개를 제외하여 조성한 후 반응을 확인한 결과 4개의 primer 들이 포함된 positive control에서만 증폭이 확인되었다. 이는 SMV의 RT-LAMP 반응에서 4개의 primer들이 모두 있어야 함을 나타낸다(Fig. 5).
Fig. 3
Specific reaction of primers for target virus. Lane M, 100 bp DNA ladder; lane 1, SMV; lane 2, SYMMV; lane 3, SYCMV; lane 4, PSV; lane 5, BCMV; lane 6, PeMoV; lane 7, negative control.
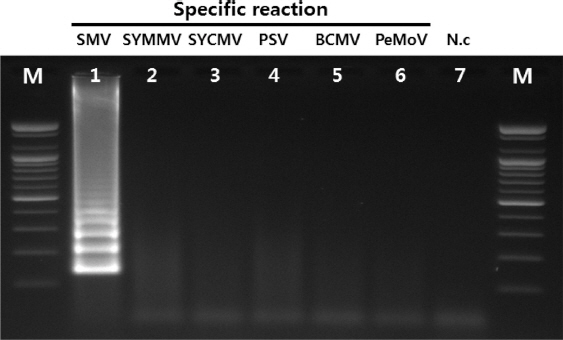
Fig. 4
Specific reaction of primers for nine Soybean mosaic virus strains. Lane M, 100 bp DNA ladder; lanes 1-9 (SMV strain G1, G2, G3, G5, G5H, G6, G6H, G7 and G7H, respectively); lane 10, negative control.
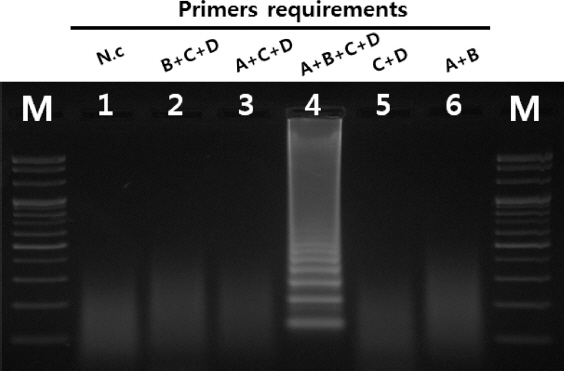
Fig. 5
Requirements for four LAMP primers. Lane M, 100 bp DNA ladder; lane 1, negative control; lane 2, B+C+D; lane 3, A+C+D; lane 4, A+B+C+D lane 5, C+D; lane 6, A+B(A: FIP, B: BIP, C: F3, D: B3).
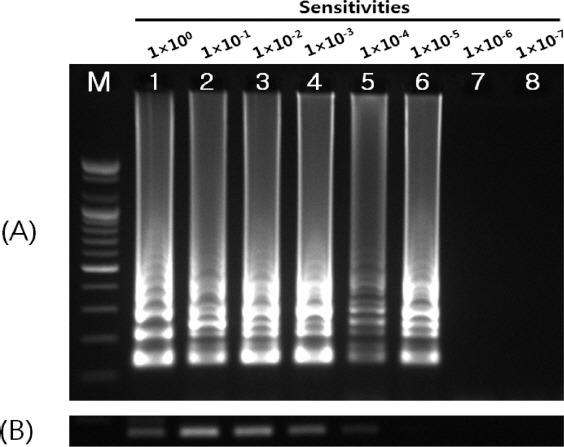
RT-LAMP는 total RNA를 주형으로 사용하는 것으로 전체 RNA내에서 특정 mRNA의 농도를 확인하기 위하여, SMV의 RTLAMP로 증폭되는 부위를 포함하는 DNA clone을 주형으로 하여 민감도를 확인하였다. DNA clone의 product size는 295 bp 이며, 확인된 정량은 147 ng/ul이었다. 이 DNA clone을 주형으로 1×100부터 1×10-7까지 단계적으로 희석하여 RT-LAMP와 RT-PCR을 수행한 결과 RT-LAMP는 1×10-5까지 검출이 가능한 것으로 확인되었다(Fig. 6A). RT-PCR의 검출 한계는 1×10-4 정도로 RT-LAMP의 민감도가 10배 높은 것으로 확인되었다 (Fig. 6B).
Fig. 6
Sensitivities of (A) RT-LAMP and (B) RT-PCR for detection of Soybean mosaic virus. LAMP was performed under 58oC and 60 min. Lane M, 100 bp DNA ladder; lanes 1-8 (1×100, 1×10-1, 1×10-2, 1×10-3, 1×10-4, 1×10-5, 1×10-6 and 1×10-7, respectively).
현장에서 RT-LAMP를 실시하기 위해서는 Thermocycler 등과 같은 전문기기가 아닌 간편하게 사용할 수 있는 기기가 필요하다. 따라서 항온수조와 heating block과 같은 간편한 등온 장치에서 재현성을 확인하기 위해 Thermocycler 기기와 비교 하여 반응 여부를 확인하였다. 세 가지 기기에서 58°C, 60분 동안 실시된 반응을 전기영동으로 확인한 결과 모두 동일한 반응 결과를 나타내었다(Fig. 7). 또한, SYBR Green I을 이용하여 반응 tube내에서 색에 의한 반응 여부를 확인하였다. 색의 변화는 자연광, portable UV light와 UV transilluminator에서 그 반응을 확인하였다. 자연광에서도 그 증폭여부의 확인이 충분히 가능 하나 portable UV light와 UV transilluminator를 이용할 경우 확실한 반응여부를 확인할 수 있었다(Fig. 8). 하지만 현장에 적용 하기 위해서는 viral RNA의 간편한 추출과 어디서든 사용 가능한 등온 장치의 이용이 고려되어야 할 것이다.
Fig. 7
Specific reaction of primers depends on using machine. The total RNA from infected plant by SMV and healthy plant were used as a positive and negative control, respectively. Lane M, 100 bp DNA ladder; lanes 1-2 (positive and negative control in Thermocycler machine); lanes 3-4 (positive and negative control in water bath); lanes 5-6 (positive and negative control in heating block).

Fig. 8
Fluorescent dye mediated visual detection for RT-LAMP. The total RNA from infected plant by SMV and healthy plant were used as a positive and negative control, respectively. A: Fluorescent detection from positive (white, left) and negative (colorless, right) under UV transilluminator; B: Fluorescent detection from positive (green, left) and negative (colorless, right) under portable UV light; C: Naked-eye detection from positive (green, left) and negative (yellow, right) under normal light.

 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






