

벼 도열병은 벼 및 화본과 작물에 병을 일으키는 병원균인 Magnaporthe oryzae에 의해 발생된다. 전 세계적으로 벼를 재배하는 지역에서 가장 중요한 병으로 벼의 유묘기에서 수확기에 이르기까지 벼의 전 생육기에 걸쳐 나타나며, 이 병에 의한 손실량은 한 해 6000만 명이 먹을 수 있을 정도에 달한다(Ou, 1985). 우리나라의 경우 1970년대 벼의 생산량 증대를 위해 개발된 통일계 품종들이 전국적으로 재배되고, 기주의 단일화와 환경조건이 병원균 발생에 유리한 상황으로 지속되면서 벼 도열병이 대발생된 사례가 있다. 또한 최근 이상 기온 현상으로 2010년 미국에서 벼를 가장 많이 재배하는 Arkansas 지역에서 벼 도열병이 대발생하여 큰 피해를 초래했으며, 인도네시아에 서는 2010년과 2011년 각각 10,604 ha와 11,929 ha가 손실을 입었다(Wibowo, 2011).
도열병균은 벼의 그루터기 등에서 균사체나 포자 상태로 월동하는 것으로 생각되고 있으며, 벼가 논에 이식되어 재배되 기 시작하는 시점의 유묘기 때부터 잎에 침입하기 시작한다. 다른 식물 병원성 곰팡이와 마찬가지로 벼 도열병균의 무성포자가 가장 중요한 최초의 감염체로, 무성포자는 기주 부착 후 1-;2시간 내 발아가 시작된다(Bourett와 Howard, 1990; Hamer 등, 1988). 발아된 포자는 대략 6-8시간 정도 지나면 침입구조인 부착기(appressorium)를 형성하는데, 이 부착기는 세포내 glycerol의 축척에 의해 8 MPa이라는 엄청난 압력으로 단단한 기주 식물을 뚫고 침입한다. 이러한 일련의 과정은 24-;36시간 내에 이루어진다(Bourett와 Howard, 1990; Hamer 등, 1988; Khang 등, 2010; Mosquera 등, 2009). 한 개의 병반에서 하루 밤 사이 2000-6000개의 새로운 무성포자를 생산하여 다른 기주로 빠르게 옮겨갈 수 있으며(Oh와 Ayad,1967), 짧게는 4일에서 일주일내에 한 세대의 생활사를 완성한다(Oh와 Ayad, 1967). Han 등(1997)에 의한 생태학적 조사에 의하면 우리나라에서는 벼의 재배기간 동안 8-12회의 생활사를 완성하는 것이 밝혀졌다. 대발생 피해는 주로 벼의 수확기에 발생하는 목 도열병 및 이삭 도열병에 의해 야기된다. 이삭이 맺히는 초기에 목 도열병이 발생할 경우 이삭이 정상적으로 영양분을 공급받지 못해 알곡이 가득 차지 않아 이삭이 말라버리거나 떨어지게 되며, 이삭에 직접 병징이 발생할 경우 이삭의 발달이 정상적으로 이루어지지 않기 때문에 심각한 수량 손실이 발생하게 된다(Agrios, 1997).
도열병균의 병원성 검정은 일반적으로 3-4엽의 유묘기 벼를 이용하여 수행된다(Valent 등, 1991). 연구자들 간의 약간씩의 차이가 있지만 일반적인 접종 방법은 다음과 같다. 3-4엽기의 벼에 적정량으로 희석한 무성포자 용액을 접종하며, 접종 이후 에는 빛이 들지 않는 암조건의 고습도생장실에 16-24시간 배양한다. 이후 상온 조건의 식물 생장상으로 옮겨 약 7일간 병반 발생을 관찰한다.
벼의 잎을 이용한 병원성 검정법은 도열병균의 병원력을 손쉽게 검정하는데 매우 유리한 방법이지만, 목 도열병 또는 이삭 도열병과 같이 다른 조직에서 분리된 도열병균의 병원력을 검정하거나 혹은 도열병균이 조직특이성을 갖는지의 여부를 판단하기에는 적합하지 않다. 특히, 대부분의 수량 손실을 불러오는 도열병이 목 및 이삭 도열병임을 감안하면 유묘기의 잎을 활용한 병원성 검정법과 함께 목 및 이삭기의 벼를 이용한 도열병검정법이 요구된다. 그러나 아직까지 목 및 이삭 도열병의 검정을 위한 실험 방법은 개발되지 않았다. 이 연구에서는 이삭기에 접어든 4개월 된 성체 식물을 이용하여 다른 조직에서 분리된 대표균주를 이용하여 벼 도열병균의 접종법을 개발하였다.
대표균주로서 선발된 균주는 모두 서울대학교 유전자원센터에 장기 보존된 균주를 분양받아 사용하였다(Table 1). 도열 병 대표균주는 1999년 유묘기부터 이삭기까지의 벼 품종 추청에 발생한 병반으로부터 분리되었다. 이들은 각각 잎(F1-7), 목(Fcn12-17), 이삭(Fcn14-1)에 발생한 병반에서 분리된 균주로서 경기도농업기술원(경기도 화성시 기산동 315-1)의 동일한 시험포장에서 각기 다른 생육기에 분리되었다(Table 1).
Table 1
Three representative isolates collected from Oryza sativa cv. Chuchungbyeo at Kyunggi-do Agricultural Research and Extension Services (Hwaseong, Korea) in 1999 and used in development of virulence test methods for neck and panicle blast
| Isolate | Lesion | Collection date |
|---|---|---|
| F1-7 | Leaf blast | June 30 |
| Fcn12-17 | Neck blast | September 15 |
| Fcn14-1 | Panicle blast | September 29 |
기주 식물로써 도열병 대표 균주가 분리되었던 ‘추청벼’ (Oryza sativa cv. Chuchungbyeo)를 실험에 사용하였다. 종자 처리는 95% 에틸알코올에 1분간 침지한 뒤 1% 하이포아염소산나트륨 수용액에 1분간 재 침지하고, 이후 하이포아염소산나트륨을 제거하기 위해 10회 이상 증류수로 씻어주었다. 발아를 위해 25°C 생장상에서 3-4일간 배양하며 하루에 한번 물을 갈아주었다. 잎 도열병 검정에 사용한 유묘는 벼 못자리용 육묘상토(질소전량 800-2,500 mg/kg, 유효인산 150-160 mg/l, EC 2.0 dS/m)가 담긴 5 × 5 × 5 cm의 플라스틱 포트에 포트 당 4립씩 파종하고, 25°C 식물 생장상에서 명/암 조건이 16/8시간 상태로 약 3주간 육묘하였다. 목 도열병 및 이삭 도열병 검정을 위해 사용된 벼는 경기도농업기술원의 포장에서 4개월간 자란 ‘추청벼’ 중 병징이 없는 성체 식물을 선발하여 대형 포트에 옮겨 심은 뒤 실험에 사용하였다.
선택된 균주의 포자형성을 위해 귀리한천배지(Oatmeal Agar medium: 귀리 50 g, 한천 20 g, 증류수 1 l)와 V8-주스한천배지(V8-Juice 80 ml, 한천 15 g, 증류수 920 ml, pH 6.8)를 사용하였 다. 배지는 고압멸균한 후 직경 9 cm Petri-dish에 분주하여 굳혀 준비하였다. 도열병 균총의 절편을 귀리한천배지 또는 V8- 주스한천배지에 접종 한 후 지속적으로 빛이 조사되는 25°C 항온기내에서 7-14일간 배양하여 무성포자를 형성시켰다.
접종원의 준비는 무성포자가 형성된 Petri-dish에 Tween 20를 1000배로 희석한 멸균 증류수를 10 ml 부은 뒤, 멸균된 스프 레더를 이용하여 포자를 수확한 후 miracloth (Calbiochem, San Diego, CA)로 균사를 걸러내 무성포자 현탁액을 제조하였다. 포자 농도를 hemacytometer로 측정한 뒤 1 × 105/ml로 농도를 조정하여 접종원을 준비하였다(Fig. 1).
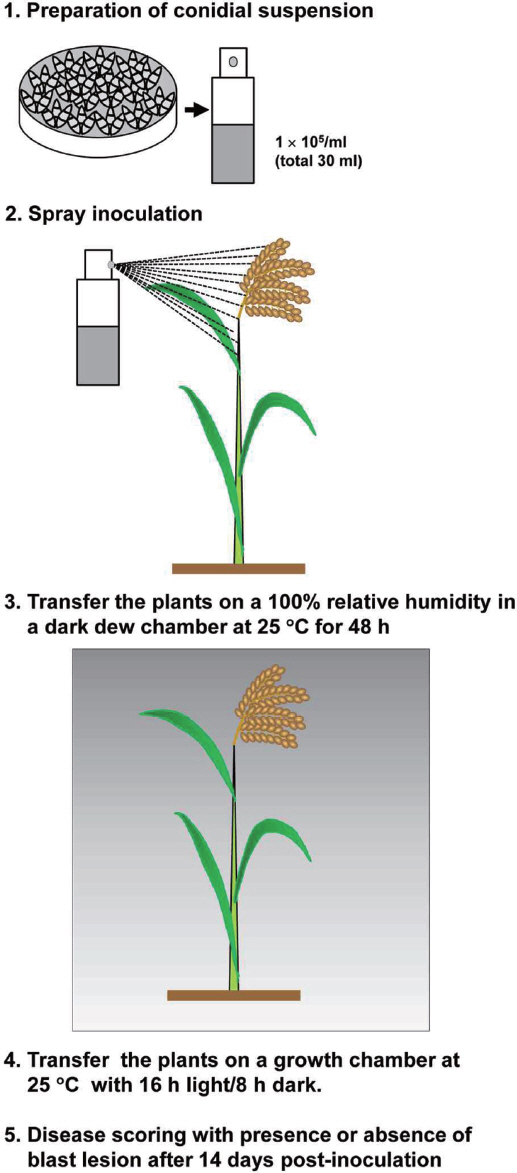
병원성 검정은 기본적으로 기존의 유묘 식물체를 이용한 방법을 따랐으나, 벼 도열병의 대발생에는 장기간의 저온 다습한 환경조건이 수반되는 점에 착안하여 성체 식물 접종 시 습실 배양 기간을 다르게 적용하였다. 포자 현탁액의 접종은 1회용 스프레이어를 이용하여 이루어졌으며 유묘 식물체 크기에 비해 이삭기에 접어든 성체 식물은 3배 이상 표면적이 넓기 때문에, 유묘 접종 시 한 포트 당 10 ml, 그리고 목 및 이삭에 접종 시 포트 당 30 ml의 포자현탁액을 분무하여 접종하였다. 접종 된 식물은 25°C, 암조건의 고습도생장상에 넣어 유묘의 경우 24시간, 성체 식물의 경우 24시간 또는 48시간 습실 처리 하였다. 이후 식물 생장상으로 옮겨 명/암 조건이 16/8시간 상태로 유묘의 경우 7일간, 성체 식물의 경우 14일간 배양하면서 매일 병반 발생 여부를 확인하였다(Fig. 1).
발병조사는 유묘의 경우 식물체 전체 잎에 형성된 병반형 및 병반수를 계수하여 Valent 등(1991)에 의한 발병지수에 의해 0-5로 기록하였다: 0-2는 비병원성; 3은 중도병원성; 4-5는 고도병원성으로 구분하여 단계별로 조사하였다. 목과 이삭의 경 우 병반 면적률은 크게 의미를 가지지 않기 때문에 발병 정도를 감염 개체율로 계산하였다. 계산은 접종한 목 및 이삭의 총 개체 수에서 감염된 개체수를 계수하여 백분율로 환산하였다.
실험에 선발된 3개의 도열병 균주는 5개 이상의 ‘추청벼’의 유묘에 평균 발병지수 4이상의 고도병원성을 보였으며, 균주간 의 병원성의 차이가 통계학적으로 없었다(Fig. 2A). 발병지수 4 이상은 벼 잎에 대략 3-4 mm 길이의 안점(eyespot) 병반이 다수 보이는 경우였다(Valent 등, 1991). 이 결과는 우리가 실험 에 사용하기 위해 선발한 3개의 균주 모두 최초 분리된 조직의 종류에 관계없이 유묘에 대해 병원력을 가지고 있음을 나 타내었다.
Fig. 2
Pathogenicity assay. (A) Pathogenicity assay on rice leaves. Ten milliliters of conidial suspension (1 × 105 conidia/ml) of each isolates were sprayed on 3-4 leaf stage of rice seedlings (Oryza sativa cv. Chuchungbyeo). The inoculated plants were kept in a dew chamber at 25°C for 24 h in darkness and moved to a growth chamber with a photoperiod of 16 h with fluorescent lights. Scores of disease severity were measured 7 days after inoculation, as previously described (Valent et al., 1991). (B) Pathogenicity assay on rice neck and (C) on rice panicles. Thirty milliliters of conidial suspension (1 × 105 conidia/ml) of each isolates were sprayed on panicle stage of rice (Oryza sativa cv. Chuchungbyeo). The inoculated plants were kept in a dew chamber at 25°C for 24 h or 48 h in darkness and moved to a growth chamber with a photoperiod of 16 h with fluorescent lights. Scores of diseased neck and panicles were counted 14 days after inoculation. Tukey’s test was used to determine significance at the 95% probability level. The same letters on the standard deviation showed no significant difference.
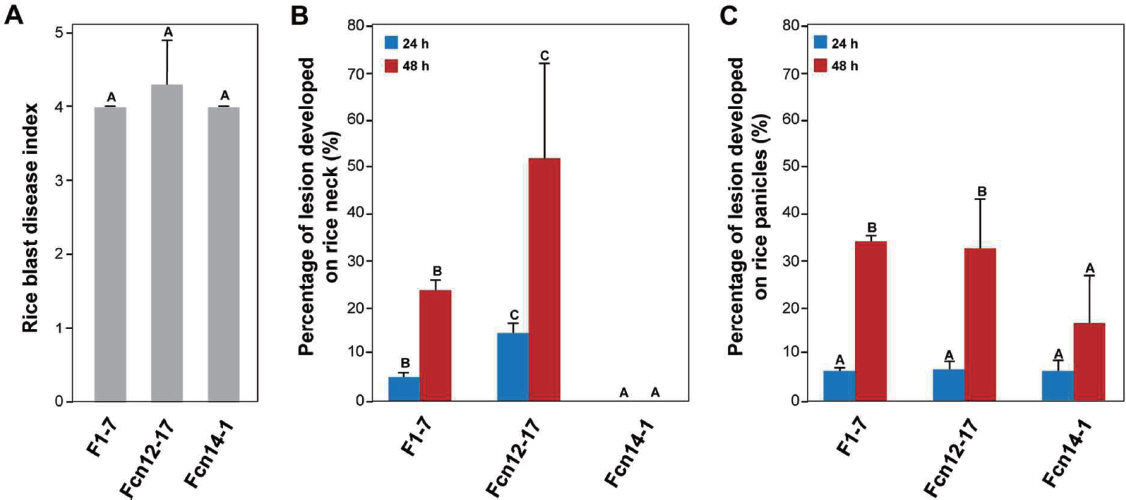
성체식물 접종은 3개 포트의 독립적인 식물체를 한 균주 접종에 사용하였다. 하나의 포트 내에 대략 14-20개의 목과 이삭 이 관찰되었으므로, 대략 50-60개 정도의 목과 이삭이 한 균주에 대한 접종 실험에 사용되었다. 실험 처리 간 비교를 위하여 통계학적인 분석방법은 Tukey’s HSD를 사용하였다. 선발된 도열병균을 각 실험구당 3개 이상의 성체 식물에 접종했을 때, 우리가 예상한 바와 같이 오랜 시간(48시간) 습실 처리 시, 좀 더 명확한 병징을 관찰할 수 있었다. 24시간 습실 처리 조건에서는 48시간 습실 처리 조건에 비해 매우 낮은 병 발생율(5-10%)을 보였기 때문에, 24시간 습실 처리 조건은 충분한 병징을 발생시 키기에 부족한 조건으로 판단되었다(Fig. 2B, Fig. 2C). 또한 유묘의 경우 접종 후 7일 뒤 병원성 여부를 판별하기에 충분한 병징이 관찰되는데 반해 성체 식물의 경우 접종 후 14일이 되어서야 충분한 병징이 관찰되었다. 이러한 결과는 목 도열병 및 이삭 도열병 검정을 위한 접종 실험시 적어도 48시간 이상의 습실 처리와 2주 이상의 병반 형성 기간이 필요함을 나타낸다.
흥미롭게도, 선발된 도열병균을 성체 식물의 목과 이삭에 접종했을 때에는 유묘에서와는 달리 세 균주의 병원성이 다르게 나타났다. 특히, 이삭에서 분리된 Fcn14-1은 벼의 목에 전혀 병을 내지 못했다. 반면에, 잎에서 분리된 F1-7이 23.7%로 상대적으로 낮은 발병률을 보였으며, 목에서 분리된 Fcn12-17은 51.5%의 높은 발병률을 보였다(Fig. 2B). 이삭의 경우 모든 균주에서 15% 이상의 발병률을 보였다(Fig. 2C). 특히 잎에서 분리된 F1-7과 목에서 분리된 Fcn12-17은 이삭에서 분리된 Fcn14-1에 비해 높은 발병률(>30%)을 보였다. 이러한 결과는 식물의 조직과 병원균의 종류에 따라 적합성(compatibility)이 달라질 수 있다는 것을 나타낸다. 실제 우리나라의 포장에서 잎 도열병과 이삭 도열병의 발병률이 크게 차이가 난다는 보고가 있었는데 (Ra 등, 1993; Ra 등, 1994), 이것은 잎 도열병과 이삭도열병 균주 간의 병원력에 차이가 있을 수 있다는 가능성을 제시하고 있다. 본 연구에서 개발된 목 및 이삭 도열병 검정 방법은 기주인 벼의 유묘기부터 이삭기에 따른 도열병균의 포장 내에서의 변이 및 조직-특이성을 판별하는데 도움을 줄 것으로 기대한다.
상기한 바와 같이, 본 연구에서는 성체 식물을 이용한 목 도열병 및 이삭 도열병 검정방법을 개발하였다. 특히, 잎을 이용한 벼 도열병 검정법과 다르게 성체 식물을 이용한 벼 도열병 검정법은 48시간 이상의 습실처리가 요구되며, 병반 형성에 2주 이상이 소요됨을 보였다. 이 방법은 포장 내에서 벼 도열병균 집단이 어떻게 변이하는지를 병원력 수준에서 밝히는데 활용 될 수 있을 뿐 아니라 벼 도열병 균주들이 기주의 조직에 따라 어떻게 적합성이 달라지는지를 밝혀내는데 사용 가능할 것이다.
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






