



Introduction
Tomato (Solanum lycopersicum) is one of the most important crops in the world's diet (Friedman, 2002; George et al., 2004). Tomato plants are widely grown worldwide (a total of 4,762,457 ha) and their fruits are consumed in large quantities in the traditional diet. In Palestine, tomato plants are grown all year round either in greenhouses or in open fields. The total cultivated area was 1,543 ha in 2018 with an average annual production of 220,864 tons (Food and Agriculture Organization of the United Nations, 2020).
Viruses are the most common cause of diseases affecting plants around the world and are important limiting factors in many crop production systems. These diseases result in losses through the reduction in growth, yield, and malformation of fruits. Different tomato plant varieties are grown in protected greenhouses and net-houses are highly exposed to infections by mechanically transmitted viruses or viroids primarily by the prevalent genera of the Pospiviroids, Potex-viruses, and Tobamoviruses (Luria et al., 2017).
According to the 2020 release of the International Committee on Taxonomy of Viruses (ICTV, http://www.ictvonline.org/virustaxonomy.asp), the Tobamovirus genus includes the largest number of species (37 species) among the seven genera in the family Virgaviridae. The Tobamovirus genus includes the well-known species: Tobacco mosaic virus (TMV) (Mayer et al., 2009), Tomato mosaic virus (ToMV), Tobacco mild green mosaic virus (TMGMV), Tomato mottle mosaic virus (ToMMV) as well as Pepper mild mottle virus (PMMoV) among the viruses capable of infecting Solanaceae crops (Adams et al., 2009; Ambros et al., 2017; King et al., 2012). Tobamoviruses are characterized by a 300-nm-long rod-shaped particle encapsulating a positive single-stranded RNA genome of 6.2 to 6.4 kb encoding four proteins. The genomic segment expresses two proteins of 126 and 183 kDa, resulting from partial suppression of a stop codon and encoding non-structural proteins that form the replicase complex; a 30-kDa movement protein (MP) is expressed through a subgenomic RNA1; and a 17.5-kDa coat protein (CP) is expressed from a second subgenomic RNA2 (Adams et al., 2009).
Tobamoviruses are easily transmitted mechanically during the cultivation process, they have no vectors in nature, are highly stable, and can remain viable in the soil for long periods in crop residues, therefore, they are capable of infecting newly established plants through possible roots injuries, which are common during root growth and expansion in the soil (Candemir et al. 2012; G├╝lser et al. 2008).
The frequent evolution or outburst of new viruses or isolates of already known viruses reduces the annual yield and quality of tomato production. A new tobamovirus species, the Tomato brown rugose fruit virus (ToBRFV), has been characterized by infecting tomatoes in Jordan (Salem et al., 2016), Israel (Luria et al., 2017), Northern West Bank (State of Palestine, SP) (AlKowni et al., 2019), Germany (Menzel et al., 2019), Turkey (Fidan et al., 2019), Mexico (Camacho-Beltr├Īn et al., 2019), the United Kingdom (Skelton et al., 2019), the United States (Ling et al., 2019), Italy (Panno et al., 2019), and China (Yan et al., 2021). Tobamoviruses are transmitted mechanically through workersŌĆÖ hands, clothes, and tools, and are capable to preserve infectivity in seeds and contaminated soil (Broadbent, 1976; Dornai et al., 1993).
During the fall of 2016 and 2017, virus-like symptoms consisting of mosaics on leaves with occasional leaf narrowing, and yellow-spotted fruits were observed on tomato (Solanum lycopersicum L.) cv Ikram plants grown in greenhouses in several tomato cultivating areas of the West Bank (Jenin, Qalqilia, Tulkarm, and Nablus). The current study documented the symptoms and host range of ToBRFV isolate from the State of Palestine (ToBRFV-PAL), checked for the resistance of tomato cultivars commercially available in Palestine, and conducted identity and phylogenetic analysis of ToBRFV-PAL with another tomato- infecting tobamoviruses.
Materials and Methods
Samples collection and RNA extraction. Leaf samples of tomato plants showing disease symptoms like mosaic leave either severe or mild, leaf narrowing, and yellow spots on the fruits, were collected to identify the etiology of the disease. Samplings were performed in the most important tomato cultivating areas in the northern part of the State of Palestine (SP) (Jenin, Nablus, Qalqilia, and Tulkarm) during the fall of 2017 and summer of 2019. To investigate the source of virus infection, leaf samples were also collected from weed species prevalent in tomato greenhouses, other crops planted in the open field close to the sampling area, and from other four tobacco fields (Nicotiana tabacum L.) in the Jenin district (Table 1). All samples were either processed directly or stored at -80┬░ C for further analysis.
Table┬Ā1.
Plant species collected from cucurbit fields and tested for ToBRFV infection by RT-PCR using the specific primer pair
For RNA extraction, leaf samples were ground under liquid nitrogen. RNA was extracted from the ground samples using Qiagen RNeasy Plant mini kit (Qiagen GmbH, Hilden, Germany) following the manufacturer's instructions, and samples were stored at ŌłÆ80┬░ C until used. DNA was isolated using the CTAB (cetyltrimethylammonium bromide) method (Doyle, 1991).
Reverse transcription polymerase chain reaction and sequencing. The viral RNAs were reverse-transcribed using random hexamer primers (Roche Diagnostic, Munich, Germany) and Enhanced Avian HS reverse transcription polymerase chain reaction (RT-PCR) Kit (Sigma-Aldrich, St. Louis, MO, USA). To identify the causal agent, all samples were tested for three viruses: TMV (Kumar et al. 2011), ToMV (Kumar et al. 2011), and ToBRFV (Luria et al., 2017) that are showing similar symptoms on tomato plants using sets of primers presented in Table 2. The resulting amplicons from the ToBRFV primers were cloned into the pJET1.2/blunt-cloning vector and sequenced using standard molecular techniques. Sequence homology was determined using BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi) algorithms. The two fragments show 99.51% similarity. A BLAST search against the NCBI NR database revealed that one of the sequenced fragments (MK881101) was 99.51% identical to a region of the replicas protein (RP) gene of the ToBRFV isolate Israel /ToBRFV IL (GenBank KX619418) in the genomic RNA (gRNA) positions 3501 to 4922. The second isolate (MK881102) showed 99.72% identity with the Israeli isolate.
Table┬Ā2.
List of primers used in this study for the detection of tomato viruses, and for the cloning of ToBRFV
Using the sequence data of the ToBRFV Israeli isolate (KX619418.1), six specific primers pairs were used to amplify overlapping fragments of the full-length ToBRFV Palestinian genome. Reverse transcription was performed using random hexamer primers as mentioned previously. The resulting cDNA was amplified by polymerase chain reaction (PCR) using Phusion high-fidelity DNA polymerase (Thermo Fisher Scientific Inc., Austin TX, USA) and the sets of primers described in Table 2, and analyzed on 1.0% agarose gels in 1├Ś TAE buffer (Sambrook et al., 1989), and visualized by UV illumination using the GelRed and Gel Documentation System (U: Genus, Syngene, Cambridge, UK). The ToBRFV cDNA products were cloned into the pJET1.2/blunt-cloning vector. Two PCR amplicons for each fragment were sequenced bidirectionally using forward and reverse virus-specific primers (Table 2) (Macrogen Inc., Seoul, Korea). The sequences obtained were assembled using CLC main workbench 5.6 software to obtain the full-length sequence of ToBRFV of the Palestinian isolate. The names, sequences, and locations of primers used for the amplification of ToBRFV are listed in Table 2.
Virus inoculation and host range analysis. Purified viral particles served for mechanical inoculation of commercial tomato plants. Symptomatic tomato plants collected from the field served for sap-mechanical inoculation assays. infected leaves and fruits were ground in 0.01 M sodium phosphate buffer (pH 7.0; 1:2 w/v), and the extract was rubbed on a range of commercial tomato varieties (pre-dusted with carborundum) cultivated by the Palestinian farmers, was performed with five replicates for each variety with negative control. Inoculated plants were grown at 24┬░ C in a growth chamber, and symptoms were evaluated 10 to 20 days post-inoculation. At 20 days post-inoculation, the presence of To-BRFV in the newly developed leaves was determined by RT-PCR detection.
Sequence identity and phylogenetic tree analyses. To-BRFV-PAL sequences were aligned by using CLUSTALX as implemented in MEGA6.06 (Tamura et al., 2013). phylogenetic trees were constructed using the maximum likelihood, neighbor joining, and minimum evolution methods that are packaged in MEGA version 10.1.5 (Kumar et al. 2018). Nodes with bootstrap values less than 50% were deleted, and those with bootstrap values higher than 70% were kept for 1,000 replicates.
Results
Field survey and epidemiology of the disease. In fall 2017, and summer 2019 leaf samples were collected from 100 tomato plants, distributed on 18 tomato fields in the northern West Bank, SP, that showed disease symptoms similar to those previously described to be caused by tobamoviruses. Infected plants showing symptoms including a combination of symptoms were induced by ToRBFV on tomato plants in the field. These include varying degrees of mosaic, narrowing and rolling of the leaves, and brown necrosis on the pedicles and the sepals. A typical symptom of brown rugose necrotic lesions on fruits, in addition to yellow spots on fruits of some tomato cultivars. Fruit symptoms reduced the commercial value of tomatoes (Fig. 1).
Fig.┬Ā1.
ToBRFV-infected tomato (Solanum lycopersicum) plants. (A) Narrowing accompanied by mottling leaves. (B) Necrotic symptoms on pedicle (stem), calyces, and petioles. (C, H) Mosaic pattern developed on leaves. (D) Blisters on fruits. (E) Brown rugose symptoms developed on fruits. (F, G) Chlorotic spots on fruits. ToBRFV, Tomato brown rugose fruit virus.

Disease incidence (number of symptomatic plants/50 plants) reached 100% in some of the surveyed tomato fields. Analysis of the collected samples by RT-PCR using specific primers revealed that 78% (78 out of 100) of the samples were infected with tobamoviruses (Table 3). Leaf samples collected in 2017 from Jenin and Nablus were found to be mixed infected with ToBRFV and TMV (100%) (Table 3). However, tomato leaf samples collected from Tulkarm and Qalqilia have shown to be infected with ToBRFV only with an infection rate of 76% and 72.5%, respectively.
Table┬Ā3.
Detection of ToBRFV and TMV in tomato fields in Palestine during 2017-2019 using RT-PCR
Host range determination. A partial host range analysis of the newly emerging Palestinian ToBRFV was carried out. Leaf samples of symptomatic and symptomless weed species prevalent in tomato greenhouses, and from other five nearby tobacco fields in Jenin Governorate were collected and examined for the presence of ToBRFV, ToMV, and TMV using RT-PCR (Table 1). ToBRFV was detected in the following symptomless plants: Erigeron bonariensis, Chenopodium murale, Solanum melongena, and Capsicum annum (Bell pepper), and ToBRFV in RT-PCR detection. However, Chrozophora obliqua, Orobanche aegyptiaca, Portulaca oleracea, and Prosopis farcta were symptomless and negative in RT-PCR. On the other hand, Nicotiana tabacum plants were symptomatic and showed positive reactions for ToBRFV and TMV in RT-PCR reactions. N. tobaccum plants exhibited different symptoms including yellowing and different degrees of mosaic.
Biological characterization of the new Palestine/ToBRFV. To undergo the biological characterization of the new Palestine/ToBRFV isolate, we used sap extracts from the ToBRFV-PAL-infected tomato source to inoculate new S. lycopersicum plants of 17 different commercial tomato varieties collected from seed companies and Palestinian farmers in the the the study area (Table 4). Two weeks later, the inoculated tomato plants developed mosaic symptoms similar to the source plants. The following specific symptoms were observed: yellowing, curling, rolling, narrowing, or mosaic (Fig. 2). To confirm the presence of the virus in some of the inoculated plants, a one-step RT-PCR method was employed, using F-3666 and R-4718 specific primers (Table 2). ToBRFV-PAL could be detected in all seedlings of the 17 cultivars by RT-PCR.
Fig.┬Ā2.
Commercial tomato cultivars inoculated with ToBRFV-PAL showing disease symptoms on younger leaves: (A) mosaic formation on the younger leaves, (B) narrowing and rolling of leaves, (C, D) deformation of the younger leaves which may also be crumpled or puckered. ToBRFV-PAL, Tomato brown rugose fruit virus isolate from the State of Palestine.

Table┬Ā4.
A new tobamovirus isolate infects commercial tomato cultivars used by Palestinian farmers
Genome organization of ToBRFV-PAL. The complete genomic sequence of ToBRFV-PAL contained 6,392 nt (GenBank accession nos. MN013187/F42-PAL isolate and MN013188/F48-PAL isolate) with four open reading frames (ORFs) (Fig. 2). The 5ŌĆ▓- and 3ŌĆ▓-untranslated regions consisted of 74 and 201 nt, respectively. ORF1 and ORF2 encoded 126, and 183-kDa proteins, both are replicase proteins. ORF3 encoded the 30 kDa MP, and ORF4 encoded the 17.5 kDa CP (Fig. 3).
Fig.┬Ā3.
Schematic presentation of genome organization and sequencing strategy for retrieving the tobamovirus isolate genome sequence. (A) Schematic diagram of genome organization showing the viral four predicted ORFs. The numbers at the borders of each ORF represent the nucleotide base position of the start and termination codons of each ORF. (B) Selected reverse transcription polymerase chain reaction and primer sets used to amplify the complete viral genome. UTR, untranslated region; RdRP, RNA-dependent RNA polymerase; MP, movement protein; CP, coat protein; ORF, open reading frame.
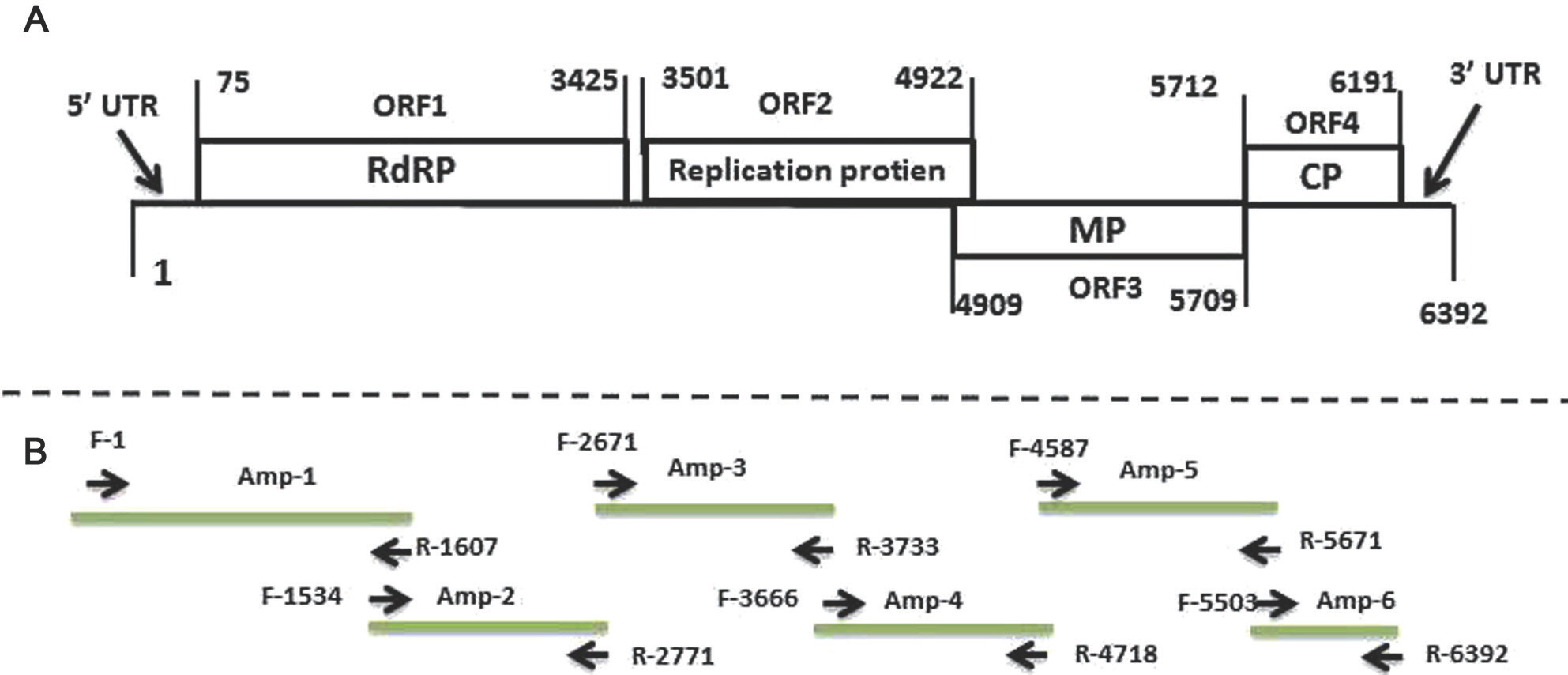
Sequence comparison and analysis of ToBRFV-PAL. To obtain the full genomic sequence of the virus, six ToBRFV-specific primers pairs were used to get an assembled nucleotide sequence through standard protocols (Table 2, Fig. 3B). The complete genome sequence of the new Palestinian To-BRFV gRNA has a length of 6,392 nt (GenBank accession nos. MN013187/F42-PAL isolate and MN013188/F48-PAL isolate) showing 18and 19 nucleotide replacements relative to the Israel/IL KX619418.1 isolate, respectively (14 transitions and four transversions in F42-PAL, while 15 transitions and four transversions in F48-PAL, leading to nine and ten nonsynonymous substitutions at the protein level in the Palestinian isolates, respectively) (Table 5). Five and nine of the replacements are substitutions affecting the RdRP gene in F42-PAL and F48-PAL, respectively (four transitions and one transversion in F42-PAL, while eight transitions and one transversion in F48-PAL) that lead to a total of four nonsynonymous substitutions at the protein level in F48-PAL (Table 5). The MP genes showed three-nucleotide replacements, all are transitions, which resulted in two nonsynonymous replacements in the MP gene (Table 5). Four and two of the replacements are substitutions affecting the CP gene in F42-PAL, and F48-PAL, respectively (two transitions and two transversions in F42-PAL, while two transitions in F48-PAL) that lead to a total of four nonsynonymous substitutions at the protein level in F42-PAL, and two nonsynonymous substitutions at the protein level in F48-PAL. Two of the transversions in the F42-PAL isolate led to nonsynonymous substitutions, mapped in the CP gene, whereas the third one did so in the RP gene; while only one transversion led to nonsynonymous substitution in RP (Table 5).
Table┬Ā5.
Nucleotide substitutions found in ToBRFV Palestine/MN013187 and MN013188 isolates regarding the Israel/IL KX619418.1 isolate
To further explore the genetic similarity between ToBRFV Palestinian strains (F42-PAL and F48-PAL), and other tobamoviruses, the entire gRNA sequences from 13 ToBRFV isolates available at GenBank, Jordan (NC_028478.1 Tom1-Jo, MK648157.1 -CaJO), Palestine (MK165457.1 UNVERI-FIED-Palestinian), Israel (KX619418.1), USA (MT002973.1), UK (MN182533.2), Turkey (MT107885.1), Netherlands (MN882023.1), Mexico (MK319944.1), Italy (MN167466.1), Greece (MN815773.1), Germany (MK133095.1-3H, MK133093.1-3G), and Canada (MN549394.1) were down-loaded from databases. For comparative purposes, the gRNA sequences from other tobamoviruses were also download-ed: ToMV (DQ873692.1), ToMMV (KU594507.2), and TMV (NC_001367.1). Two sequences for PMMoV (KX063611.1), and TMGMV (AB078435.1) were used as out groups.
The full-length gRNA sequences were aligned with CLUSTALX as implemented in MEGA6.06 (Tamura et al., 2013) and the genome-wide homogeneous nucleotide substitution model, GTR+G+1, was chosen for subsequent analyses. The net average nucleotide divergence between Palestine/F42-PAL and F48-PAL strains is 0.0026403┬▒0.0006639 (┬▒standard error of mean) and is 0.0026398┬▒0.0006638 between the Palestinian F48-PAL strain and the Israeli (KX619418.1) and Turkish (MT107885.1) isolates, while its 0.0028062┬▒0.0006631 with the Jordanian Tom-1 isolate, and ranged between 0.0054718┬▒0.0009746 and 0.0029714┬▒0.0007059 with other ToBRFV tested isolates. However, the net average nucleotide divergence between Palestine/F42-PAL and the other To-BRFV tested isolates ranged between 0.0033066┬▒0.0007433 for the Israeli and Turkish strains to 0.0054718┬▒0.0009746 for the Italian strain. It is worth noting that the net average nucleotide divergence between Palestine/F42-PAL, and F48-PAL strains and the UNVERIFIED-Palestinian isolate (MK165457.1) (Alkawni et al., 2019) was 0.0038┬▒0.0008 and 0.0028┬▒0.0007, respectively. In contrast, the divergence of the two Palestinian strains (Palestine/F42-PAL, and F48-PAL strains) from the other three tobamoviruses ranged between 0.2500768┬▒0.0097467 for TMV, 0.2611278┬▒0.0101505 for ToMV and 0.2629613┬▒0.0101234 for ToMMV.
A phylogenetic tree was constructed with the complete genomic sequences of the four tobamoviruses infecting tomato plants (12 accessions of ToBRFV, one accession from each of TMV, ToMV, and ToMMV) using the maximum likelihood tree, shown in Fig. 3, and its reliability evaluated with the bootstrap method (based on 1,000 pseudoreplicates). In the phylogenetic tree, viral isolates were clustered into four groups corresponding to the four tobamoviruses, in which ToBRFV and TMV formed sister branches, while ToMV and ToMMV formed another common branch (Fig. 4). It is necessary to mention that the ToBRFV isolates exhibited a ŌĆśstar phylogenyŌĆÖ pattern and shared high identities; this might be attributed to the fact that the virus has emerged recently (Yan et al., 2021).
Fig.┬Ā4.
The phylogenetic tree of ToBRFV, ToMV, TMV, and ToMMV constructed with complete genomic nucleotide sequences using the maximum likelihood method. Isolates of cucumber green mottle mosaic virus (TMGMV), and PMMoV were used as an outgroup. Nodes with bootstrap values higher than 50% were shown. To-BRFV, Tomato brown rugose fruit virus; ToMV, tomato mosaic virus; TMV, tobacco mosaic virus; ToMMV, tomato mottle mosaic virus; TMGMV, tobacco mild green mosaic virus; PMMoV, pepper mild mottle virus.

Discussion
We reported here the disease symptoms caused by To-BRFV in Palestine. Symptoms are critical in the diagnosis of plant diseases caused by viruses. ToBRFV was reported to induce brown rugose symptoms on tomato fruits in Jordan (Salem et al., 2016), in addition to yellow spots on tomato fruits in Germany (Menzel et al., 2019). It's worth noting that in Israel the virus was reported to produce yellow spots and brown rugose symptoms on cultivar Odelia (Luria et al., 2017). In the current study, ToBRFV causes several symptoms in tomato-infected plants including yellow spots on the fruits, brown rugose necrotic lesions, blistering, mosaic formation on the younger leaves which may also be crumpled or puckered, and narrowing leaves (Fig. 1). To elucidate if the symptoms were caused by a mixed infection of several viruses, we conducted RT-PCR detection of different tobamoviruses and found that the samples collected from the fields in Jenin and Nablus areas were infected with ToBRFV and TMV, while the the the samples collected from Tulkarm and Qalqilia were infected with ToBRFV only. Mixed infections with two or three viruses were also very common in tomatoes in Israel (Luria et al., 2017). ToBRFV was reported for the first time in Palestine in 2019; the disease was shown to threaten tomato production in the area, and the incidence rate was reported to reach 100% (Alkowni et al., 2019). In the current study, the disease incidence rate was close to100%, similar to what was reported previously in some fields in Jordan and Israel (Luria et al. 2017; Salem et al. 2016).
ToBRFV isolates have been reported to infect different plant species including Capsicum annumm, Datura stramonium, D. metal, Nicotiana benthamiana, N. clevelandii, N. glutinosa, N. megalosiphon, N. tabacum, Petunia hybrida, Solanum nigrum, and Chenopodium sp. (Luria et al., 2017; Salem et al., 2016). In the current study the virus has shown to infect naturally C. annum (Bell pepper), Chenopodium sp., Erigeron bonariensis, N. tabaccum, and Solanum melongena. Luria et al. (2017) have reported that ToBRFV cannot infect S. melongena, and thus suggested using grafted tomato plants on eggplant rootstock to contribute to the reduction of the primary inoculum when planting seedlings in contaminated soil, while Yan et al. (2021) suggested that ToBRFV-SD can infect S. melongena with no symptoms, however, in the current study we recorded that ToBRFV can infect S. melongena naturally, which indicate that it can't be used as rootstocks for grafting tomato.
The cultivation of N. tabaccum for commercial purposes (production of cigarettes) is quite common in the study area mainly in the Jenin Governorate. All N. tabaccum collected samples were mixed infected with ToBRFV and TMV. Tobamoviruses are known to be very stable in the environment and can survive in crops (leaves and stems) and root debris on the soil surface for at least several months and can infect a new crop planted in a contaminated site. Tobamoviruses are also carried on seeds, leading to the infection of germinating seedlings. It has been shown previously that tobacco used to make cigarettes and cigars can function as an effective disease vector, possibly helping the movement of infectious TMV between countries (Iftikhar et al., 2015). We can assume that cigarettes produced locally can be considered a source of ToBRFV transmission and infection, thus biosecurity measures should be taken among workers to limit virus transmission .
Results of identity analyses showed that ToBRFV-PAL isolates shared the highest identities, on both nucleotide and protein levels, with an isolate from Israel. Taking into consideration that the regions from which ToBRFV-PAL were isolated (Jenin, Tulkarm, and Qalqilia) are on the borders with Israel. These data suggest that ToBRFV-PAL was introduced to tomato fields in Palestine from Israel by exchange of seeds and plants between Israel and the SP, where several viruses have been disseminated worldwide through the exchange of seeds having an undetected infection (Albrechtsen, 2006; Ali-Shtayeh et al., 2014; Yan et al., 2021).
In conclusion, the current study identified a new tobamovirus isolate in Palestine that infects tomato plants. The Palestinian isolate is identical to the recently published tomato-infecting virus from Jordan and Israel. This virus is a major threat to tomato crops worldwide. ToBRFV-PAL induced mild to severe mosaic and blistering on leaves, deformation, yellow spots, and brown rugose necrotic lesions on tomato fruits. It also infected plants of C. annum, N. benthamiana, E. bonariensis, Chenopodium sp., and S. melongena. None of the seventeenth tomato cultivars tested was resistant to ToBRFV-PAL. In the phylogenetic tree constructed with the complete genomic sequences, all the ToBRFV isolates were clustered together and formed a sister branch with that of TMV. The findings of this study enhance our understanding of the biological and molecular characteristics of ToBRFV which can be helpful in the management of the disease.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






