Pectobacterium brasiliense as a Causative Agent for Soft Rot of Radish in Korea
Article information

Abstract
In October 2021, soft rot disease seriously affected radish crop in Dangjin, Chungcheongnam-do, Korea. The infected radishes were stunted and turned dark green, with yellowish leaf foliage. A slimy, wet, and decayed pith region was observed in the infected roots. The bacterial strain KNUB-03-21 was isolated from infected roots. The biochemical and morphological characteristics of the isolate were similar to those of Pectobacterium brasiliense. Phylogenetic analysis based on the sequences of the 16S rRNA region and the concatenated DNA polymerase III subunit tau (dnaX), leucine-tRNA ligase (leuS), and recombinase subunit A (recA) genes confirmed that the isolate is a novel strain of P. brasiliense. Artificial inoculation of radish with P. brasiliense KNUB-03-21 resulted in soft rot symptoms similar to those observed in infected radish in the field; subsequently, P. brasiliense KNUB-03-21 was reisolated and reidentified. To our knowledge, this is the first report of P. brasiliense as a causal pathogen of radish soft rot in Korea.
Pectobacterium species, a group of plant pathogenic bacteria, is the causative agent of soft rot disease in crop plants. Monocotyledonous and dicotyledonous plants suffering from bacterial soft rot result in serious economic loss (Ma et al., 2007). Outbreaks of blackleg and soft rot associated with Pectobacterium brasiliense have been reported in Korea (Choi and Kim, 2013; Jee et al., 2018; Park et al., 2022). Radish (Raphanus sativus) is one of the most widely grown and consumed vegetables worldwide. China is the largest radish producer in the world with planting area of 1.27 million ha and the total production of 44.6 million tons of fresh tuberous roots, accounting for 47% of global radish production (Food and Agriculture Organization, 2019). In Korea, the cultivated area of radish in 2022 was 6,340 ha, which is an increase of 422 ha (7.1%) compared with 5,919 ha in the previous year (Statistics Korea, 2022). Radishes have been cultivated continuously for decades, and they are impacted by diseases such as wilt disease, clubroot, and bacterial soft rot, which deteriorate their quality and affects yield, thereby causing heavy economic damage (Jo et al., 2011; Lee et al., 2018; Moon et al., 2001).
In September–October 2021, the radish crop was severely affected by soft rot disease in Dangjin, Chungcheongnam-do, Korea. During this period, soft rot symptoms (discoloration, decay gray, soft, wet lesions) were observed on radishes in a surveyed 25×64 m field in Dangjin. The disease incidence was found to be around 15% and the disease severity has attained to moderate level of 10.0% on radishes in the middle and later growth stages. As shown in Fig. 1A and B, the infected roots were discolored and soft rot symptoms were observed on radishes in the middle stage. In addition, soft rot symptoms were observed on the leaves of infected radishes (Fig. 1C, D). The pith region was completely decayed in infected roots in the later growth stage of radishes. Moreover, the inner part of the root contained macerated tissue and emanated a foul smell (Fig. 1E, F). However, the diseased tissue lacked visible fungal mycelia and spores. Small pieces of radish (10×10 mm) were taken, surface-sterilized for 1 min with 1% sodium hypochlorite, and then rinsed with sterile distilled water. A semiselective crystal violet pectate (CVP) medium, known as the best tool for isolation of pectolytic bacteria such as Pectobacterium spp. and Dickeya spp. (Boluk et al., 2020; Hélias et al., 2012; Oulghazi et al., 2021), was used to isolate a potential phytopathogenic agent. The collected tissues were macerated in a sterile 1.5 ml microtube with 100 μl sterile water, and approximately 10 μl suspension was spread onto CVP medium. White-gray, round bacterial colonies with entire margins and characteristic cavities in the medium, caused by their ability to metabolize pectin, mainly developed after three days of incubation at 26–28 o C. The colonies were repeatedly re-streaked onto nutrient agar plates to obtain a pure culture. As a result, bacterial strain, designated KNUB-03-21, was isolated from an infected radish.

Radish soft rot symptoms caused by Pectobacterium brasiliense KNUB-03-21 in Dangjin, Chungcheongnam-do, Korea. (A, B) The gray, soft, wet lesions were observed on the infected radish root. (C, D) The infected leaves turned dark green and showed soft rot symptoms. (E, F) The inner part of the root contained macerated tissue and emanated a foul smell. Arrowheads indicate the invasion of bacterial cells and soft rot areas.
A HiGene Genomic DNA Prep Kit (Biofact, Daejeon, Korea) was used to extract total genomic DNA from strain KNUB-03-21 for molecular analysis. The primers listed in Table 1 were used for polymerase chain reaction (PCR) amplification of the 16S rRNA region and three housekeeping genes: DNA polymerase III subunit tau (dnaX), leucine-tRNA ligase (leuS), and recombinase subunit A (recA). The 16S rRNA gene was amplified as described by Weisburg et al. (1991). A sequence of 1,352 bp was amplified from the 16S rRNA region (GenBank no. LC738895). A BLAST search of the NCBI database revealed 99.70–99.78% similarity between the 16S rRNA region sequence of KNUB-03-21 and several strains belonging to the genus Pectobacterium including P. brasiliense, P. carotovorum subsp. carotovorum, and P. versatile (Table 2). Based on these results, strain KNUB-03-21 could be recognized as a member of the genus Pectobacterium. However, the poor discriminative resolution of 16S rRNA phylogeny within this genus did not allow accurate identification of the isolate at the species level.
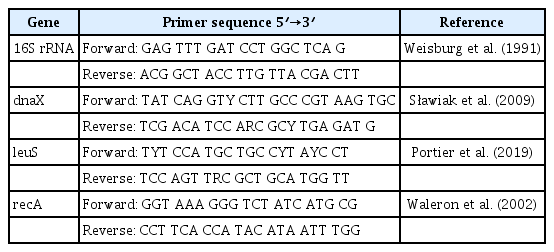
List of PCR primers used in this study
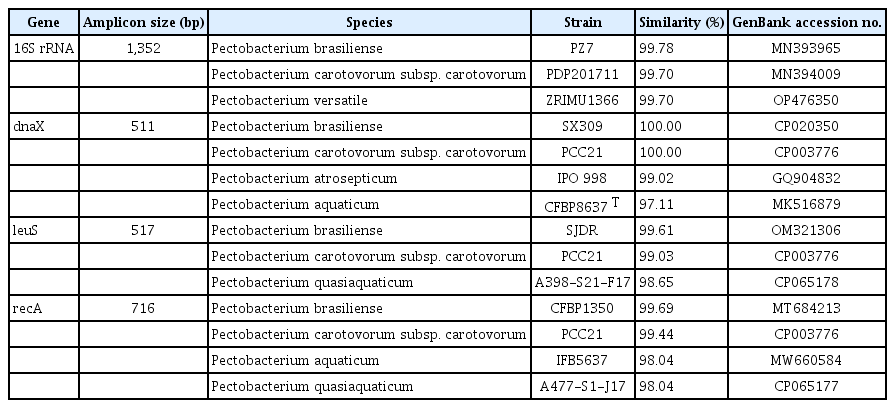
NCBI BLAST results of the 16S rRNA region sequence and amplicons generated by PCR of three housekeeping genes (dnaX, leuS, and recA) for Pectobacterium brasiliense KNUB-03-21
Recently, a phylogenetic analysis using three concatenated housekeeping genes, dnaX, leuS, and recA, allowed to update the taxonomic status of 114 Pectobacterium strains and establish P. actinidiae, P. versatile, P. odoriferum, and P. brasiliense as the novel species of the genus (Portier et al., 2019). Following this approach, dnaX, leuS, and recA genes of the strain KNUB-03-21 were amplified and sequenced. Briefly, dnaX was PCR-amplified as described by Sławiak et al. (2009) and a 511 bp sequence (GenBank no. LC738892) was obtained, which shared 100% identity with phylogenetically closely related species P. brasiliense and P. carotovorum subsp. carotovorum, 99.02% similarity with close phylogenetic relative P. atrosepticum, and 97.11% identity with close phylogenetic neighbor P. aquaticum (Table 2). The leuS gene was amplified as described by Portier et al. (2019) and a sequence of 517 bp (GenBank no. LC738894) was obtained. Based on sequence similarity, the closest relatives of strain KNUB-03-21 were P. brasiliense (99.61% similarity), P. carotovorum subsp. carotovorum (99.03%), and P. quasiaquaticum (98.65%) (Table 2). The recA gene was amplified as described by Waleron et al. (2002) and a 716 bp fragment (GenBank no. LC738893) was obtained, which showed over 98% similarity with P. brasiliense (99.69%), P. carotovorum subsp. carotovorum (99.44%), P. aquaticum, and P. quasiaquaticum (98.04%) (Table 2). Similar to the 16S rRNA phylogeny, these results indicated that the comparative analysis based on the sequence of only one of the three loci did not allow precise identification of the closest phylogenetic relatives. Therefore, phylogenetic analysis was conducted using concatenated sequences of three marker genes dnaX, leuS, and recA of strain KNUB-03-21. As mentioned above, these molecular markers have been shown to be highly effective for identifying species within the genus Pectobacterium (Portier et al., 2019). For phylogenetic analysis, the related gene sequences of the close phylogenetic relatives were obtained from the NCBI GenBank database (Table 3). The program MEGA7 was used to perform multiple sequence alignment (Kumar et al., 2016). Kimura's two-parameter model and nearest-neighbor interchange heuristic search method were used to perform the maximum-likelihood analysis and construct a phylogenetic tree (Felsenstein, 1981). A monophyletic clade composed of KNUB-03-21 and P. brasiliense strains (KNUB-01-21, CFBP5392, CFBP6607, CFBP6615, and CFBP6617 T) with a high bootstrap value strongly suggested that they belong to the same species (Fig. 2). The results of the molecular analysis showed that isolate KNUB-03-21 is a novel strain of P. brasiliense.
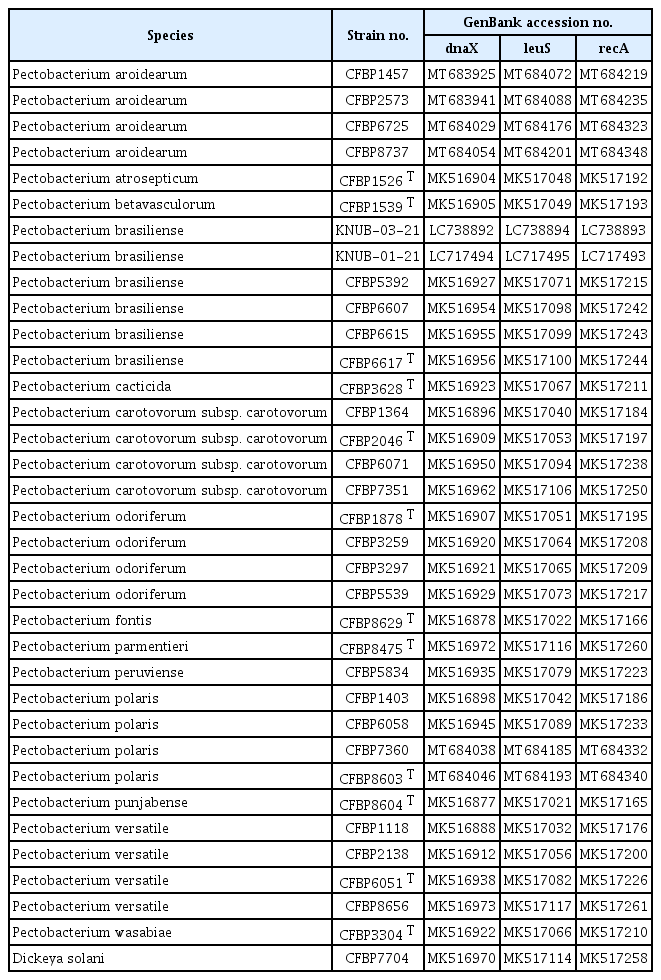
Pectobacterium species used for phylogenetic analysis and their GenBank accession numbers

Maximum-likelihood phylogenetic tree, based on concatenated partial sequences of dnaX, leuS, and recA genes, showing the phylogenetic position of strain KNUB-03-21 among related species of the genus Pectobacterium. Bootstrap values (based on 1,000 replications) greater than 70% are shown at branch points. The isolated strain is shown in bold. Dickeya solani CFBP7704 was used as the outgroup. Scale bar=0.020 substitutions per nucleotide position.
Biochemical tests were performed to confirm the identity of the isolated strain and the type strain of P. brasiliense. Strain KNUB-03-21 could grow at 37 o C, formed pits on CVP medium, showed tolerance to 5% NaCl, produced acid from α-methyl glucoside, glucose, and maltose, and formed reducing substances from sucrose. The isolate utilized acetic acid, cellobiose, D, L-lactic acid, D-melibiose, D-sorbitol, succinamic acid, and thymidine as a sole carbon source. However, KNUB-03-21 was negative for assimilation of D-arabitol, 2’-deoxyadenosine, D-glucos-aminic acid, D-glucuronic acid, glucose-1-phosphate, inosine, L-glutamic acid, maltose, Tween-40, and Tween-80 and for production of phosphatase. All these characteristics were the same to those of P. brasiliense 212 T, but several of them were different in comparison with the type species of the genus Pectobacterium, P. carotovorum subsp. carotovorum 21 T (Meng et al., 2017) (Supplementary Table 1).
Healthy radish root was used to test the pathogenicity of P. brasiliense KNUB-03-21. Briefly, the roots were washed under running tap water, air-dried, dipped in 96% ethanol, and cut into 1-cm-thick slices using a scalpel. The slices were placed in Petri dishes and their upper surface was then uniformly covered with P. brasiliense KNUB-03-21 inoculum (cell suspension: 108 cfu/ml). Rot development on slices was assessed after 48 hr incubation under greenhouse conditions (28 o C, 80% relative humidity). The pathogenicity test was repeated twice. In control group, P. brasiliense KNUB-03-21 was replaced with sterile distilled water to cover radish slices. Two days after inoculation, soft rot symptoms were observed in radish roots inoculated with KNUB-03-21. In particular, regions of completely decomposed tissue formed on the infected roots (Fig. 3A). In contrast, mock-infected radish slices showed no symptoms (Fig. 3B). The infected radish root was used to re-isolate the pathogen, and P. brasiliense was identified as the isolated bacterial strain (data not shown).

Pectobacterium brasiliense KNUB-03-21 caused radish soft rot. (A) Symptoms of soft rot induced by artificial inoculation with P. brasiliense KNUB-03-21. (B) The mock-infected radish slice showed no symptoms. All inoculated radish roots were incubated at 28 o C for 48 hr. Arrowhead indicates the invasion of bacterial cells and soft rot areas.
P. brasiliense, one of the main causative agents of soft rot, has been reported to increasingly invade various hosts worldwide. P. brasiliense causing soft rot in radish has been reported in China and is known to cause soft rot in 19 different plant species belonging to 10 different families (Liu et al., 2019; Oulghazi et al., 2021).
In this study, we identified and characterized P. brasiliense as the bacterial pathogen for soft rot recently observed in radish crop in Korea. To the best of our knowledge, this is the first study reporting P. brasiliense as the causal pathogen of radish soft rot in Korea. Our results lay a foundation for the development of control strategies to avert soft rot of plants and related economic losses caused by the identified phytopathogen.
Notes
Conflicts of Interest
No potential conflict of interest relevant to this article was reported.
Acknowledgments
This work was supported by a 2022 fund by Research of Animal and Plant Quarantine Agency, South Korea.
Electronic Supplementary Material
Supplementary materials are available at Research in Plant Disease website (http://www.online-rpd.org/).
Supplementary Table 1.
Physiological and biochemical test results of Pectobacterium brasiliense KNUB-03-21, P. brasiliense, and P. carotovorum subsp. carotovorum reference strains