



서 론
수박(Citrullus lanatus)은 전 세계적으로 약 11,720만 t이 생산되며, 한국은 세계 15위인 약 65만 t의 수박을 1만 1,972ha의 재배 면적에서 생산하고 있다. 2018년 기준, 수박은 한국인이 좋아하는 과일 선호도 조사에서 2위를 차지할 만큼 국내 소비가 많은 상황이다. 최근 수박 재배 농가들은 노지재배보다는 시설재배를 선호하며, 노동력 절감 및 높은 효율성을 위해서 대단위 공정 육묘장에서 단일 품종을 구매하여 사용하는 실정이다. 육묘장은 종자를 대량으로 파종하여 밀식 재배하고 접목하는 곳으로 작업장의 작업 도구와 극소수의 종자에 병원균이 감염되어 관리가 소홀할 경우 직접적인 피해를 받거나, 병원균이 잠복 감염될 수 있는 조건을 갖추고 있다.
수박의 과실썩음병을 유발하는 bacterial fruit blotch (BFB)는 1960년대 미국에서 처음 보고되었으며(Sowel과 Schaad, 1979), 국내에서는 1991년 전북 고창에서 처음 발생되어 현재까지도 그 피해가 보고되고 있다(Song 등, 1991). 수박 과실썩음병은 Acidovorax citrulli에 의해 발생하며, 종자 전염성 병원균으로 알려져 있으며(Hopkins와 Thompson, 2002; Rane와 Latin, 1992), 한번 발생하면 작물을 전면 폐기해야 하기에 막대한 경제적 손실을 불러오는 것으로 알려져 있다. A. citrulli는 종자나 포장에 남아 있던 식물체의 잔유물에 잠복하다 다음 해 발생하는 1차 전염 우려가 있으며(Lessl 등, 2007), 접목이나 관수 시 주변의 묘에 전염되거나, 재배 관리 작업(정식 후 적심 등)을 할 때 2차 전염의 우려가 있다(Noh와 Choi, 2015). 육묘장의 대규모 작업환경과 시설재배지의 고온 다습한 조건은 A. citrulli에 의한 과실썩음병의 발병을 유발하는 조건이 되어, 2021년 최근에도 봉화군의 수박재배지와 안성시의 육묘장에서 피해가 발생하였다.
최근 이러한 수박 과실썩음병의 피해를 방지하고자 많은 연구가 진행되었지만, 다른 식물 병원성 미생물에 비하면 미비한 상황이다. 그로 인해, 육묘장 및 농가에서 BFB가 발생하였을 시, 농약의 선택에 어려움을 겪고 있다. 최근 화학농약의 잔류 문제 및 지속성에 대한 문제로 인해 그 대안으로 생물농약에 관한 관심과 연구가 많아지고 있다.
식물 성장촉진 근권 박테리아(plant growth promoting rhizobacteria, PGPR)이란 식물 성장을 촉진하는 근권 박테리아로 최근 PGPR을 사용하여 친환경 지속 농업 개발을 위한 연구가 진행 중이다(Kumar와 Sarma, 2016). 최근 Mohammed 등(2020)은 토마토의 근권에서 분리한 Pseudomonas를 사용하여 토마토 시들음병을 유발하는 Ralstonia solanacearum의 발생을 감소시키고 실험된 토마토의 성장을 촉진하였다고 보고했다. 또한, Paenibacillus jamilae는 여러 토양 매개 병원체에 대한 높은 활성을 나타냈으며, 질소 고정, 인산염 및 칼륨 가용화, 사이드로포어 및 인돌아세트산를 생산하여 근권의 유익한 세균이 증가하는 결과를 보였다(Wang 등, 2019).
본 연구는 수박 재배지에서 분리한 A. citrulli를 억제하는 미생물을 선발하고, 선발한 미생물에 대해 각종 성장촉진 및 길항 효소 생성확인 실험을 통해, 식물 성장 촉진제 및 생물 방제제로 사용할 수 있는 잠재력을 조사하여 친환경 자재 개발 가능성을 제공하고자 수행하였다.
재료 및 방법
수박 재배지 및 수박 육묘장에서 샘플 수집 및 미생물 분리. 전국 12개 지역의 94곳의 수박재배지에서 수박 식물체(잎, 꽃, 뿌리) 및 토양을 수집하였으며, 전국 25곳의 대형 원예 육묘장에서 접목 상태의 어린 수박 모종을 수집하여 사용하였다. 수집한 수박 식물체는 각각 2 g씩 채취한 후 멸균수 5 ml 과 섞어 마쇄한 후 1 ml을 뽑아 멸균수에 희석 하였으며, 토양은 각각 1 g씩 채취하여 멸균수에 10−4까지 희석 하였다. 그 후 TSA (tryptone 15.0 g, soytone 5.0 g, sodium chloride 5.0 g, agar 15.0 g) 배지와 YPD (yeast 10.0 g, peptone 20 g, dextrose 20 g, agar 15 g) 배지를 만들어 각각 배지에 10−3과 10−4로 희석한 시료들을 30 μ l씩 도말하여 28 °C에서 24시간 배양하였다. 배지 위에 나타난 colony의 형태와 색, 크기로 육안 검정하여 서로 다른 미생물을 분리하였으며, 2-3회 반복 streaking하여 미생물을 순수 분리하였다. 순수 분리한 미생물은 다음 실험을 위해 20% glycerol stock으로 제조하여 -80 °C의 deep freezer에 보관하였다.
A. citrulli 분리 및 동정. 수박재배지 및 육묘장 샘플에서 분리된 미생물들을 한천배지 위에 배양된 colony의 모양, 색, 크기를 A. citrulli의 colony와 비교하여 육안 검정 후 A. citrulli로 의심되는 미생물의 배양액을 각각 250 μ l씩 1.5-ml centrifuge tube에 250 μ l의 멸균수와 함께 넣고 Aac Immunostrip (Agdia, Elkhat, IN, USA)을 꽂은 뒤 25-30분간 반응시켜 2개의 밴드를 보이는 미생물을 1차 선별하였다. 1차 선발한 균주의 배양액을 40 μ l씩 microtube에 분주한 후 DNA 추출을 위하여 Fast DNA kit (MPbio, Santa Ana, CA, USA)를 사용하여 추출 방법에 따라 DNA 를 추출하였다. 추출한 DNA는 A. citrulli-F: 5'-GACCAGCCAC ACTGGGAC-3', A.citrulli-R: 5'-CTGCCGTAC TCCA GCGAT-3' 프라이머를 사용하여 95 °C (5분), 95 °C (20초), 55 °C (30초), 72 °C (50초)에서 30 cycles 최종 72 °C 5분 조건에서 polymerase chain reaction (PCR)을 수행하였으며, 전기영동하여 fluorometer를 통해 밴드의 위치를 확인하였다. 선발한 균주의 동정을 위해 16S rRNA 정보를 사용하여 Macrogen (Seoul, Korea)에 의뢰하였으며, 분석기관은 Roche 454 GS-FLX+ 장비를 사용하였다. 정방향 프라이머로는 V1-9F (5′-AC-GAGTTTG ATCMTGGGCTCATCG-3′) adaptor-linker-primer의 순서로 구성되어 있으며, 역방향 프라이머로는 V3-541R (5′-X-AC-WTTACCGCGGCCTGCT GG-3′)를 이용하였다. Capillary electrophoresis sequencing을 통해 선별된 균주들을 동정하여 최종적으로 A. citrulli를 선발하였다. 선발된 A. citrulli는 NCBI의 GenBank accession 번호를 등록하였다.
선발한 A. citrulli에 대한 길항 미생물 선발. A. citrulli에 an-tagonism을 가진 세균을 선발하기 위해 50-ml 삼각플라스크에 tryptic soy broth (TSB)와 YPD를 30 ml씩 분주하여 121 °C, 15분간 고압 멸균한 후 각각 A. citrulli (HPP-3-3B, HPP-9-4B)와 전국 수박 농가의 토양 sample과 육묘장 식물체 sample에서 분리된 세균을 30 μ l 접종하여 진탕 배양기에서 28 °C, 48시간 동안 배양하였다. A. citrulli (HPP-3-3B, HPP-9-4B) 균주 현탁액(1×106 coloni forming unit [CFU]/ml)을 각각 150-mm Petri dish에 분주한 TSA 와 YPD 배지에 100 μl씩 도말한 뒤 일정한 간격으로 paper disk (ADVANTEC, Yongin, Korea)를 5장씩 올려놓은 후 액체 배양한 세균 현탁액(1×106 CFU/ml)을 25 μ l씩 접종한 뒤 항온배양기에 28 °C, 48시간 동안 배양 후 접종한 paper disk 주변의 clear zone 을 확인하여 길항 미생물을 1차 선발하였다. 1차 선발한 미생물은 90-mm Petri dish에 분주한 TSA와 YPD 배지에 A. citrulli (HPP-3-3B, HPP-9-4B) 균주 현탁액(1×106 CFU/ml)을 각각 30 μ l씩 도말한 뒤 배지 중앙에 paper disc를 올려놓고 액체 배양한 세균 현탁액(1×106 CFU/ml)을 paper disk에 25 μ l 접종한 뒤 항온배양기에 28 °C, 48시간 동안 배양하였다. 그 후, 접종한 paper disk 주변의 clear zone을 확인하여 최종적으로 A. citrulli에 길항성을 보이는 미생물을 선발하였다.
선발된 길항 미생물의 생화학적 검증.
Ammonia production: 선발된 세균을 TSB 배지에 배양하였다. 배양한 세균은 각각 50 μ l씩 30 ml의 peptone water broth (4%)에 접종하여 6일 동안 28 °C, 130 rpm으로 배양하면서 3일 간격으로 2회 조사하였다. 8.5 ml의 증류수에 2 ml의 세균 배양액, 1 ml의 Nessler's reagent (SAM, Seoul, Korea)를 혼합하여 5분 동안 반응시켜 yellow color에서 orange/red color로 변화하는 것으로, ammonia 생성 여부를 확인하였다.
Phosphate solubilization: Pikovskaya's agar (dextrose 10.0 g, calcium phosphate 5.0 g, yeast extract 0.5 g, ammonium sulphate 0.5 g, potassium chloride 0.2 g, magnesium sulphate 0.1 g, manganese sulphate 0.0001 g, ferrous sulphate 0.0001 g, agar 15.0 g/1 l)에 cork borer (⦶: 5 mm)를 이용하여 일정한 크기의 well을 만들었다. 선발된 세균을 각각 TSB 배지에 28 °C, 130 rpm, 24시간 배양한 후 20 μ l씩 well에 접종하였다. 28 °C에서 72시간 배양한 후 well 주변의 clear zone 생성 여부로 불용성 인산 가용화 능력을 확인하였다.
Starch hydrolysis: 선발된 세균을 TSB 배지에 28 °C, 130rpm, 24시간 액체 배양하였다. starch agar (soluble starch 10.0 g, peptone 5.0 g, beef extract 3.0 g, agar 15.0 g/1 l)에 cork borer (⦶: 5 mm)를 이용하여 일정한 크기의 well을 만들고, 배양된 각각의 세균을 20 μ l씩 접종하여 28 °C에서 72시간 배양하였다. 배양한 후 3% iodine-potassium iodide를 이용해 염색하여 clear zone을 확인하여 전분 가수분해 효소의 생성 여부를 확인하였다.
Siderophore production: 실험에 사용한 Chrome azurol S (CAS) 배지의 CAS blue solution은 Schwyn과 Neilands (1987)이 실험한 방법을 참고하여 준비하였다. CAS blue agar에 cork borer (⦶: 5 mm)를 이용하여 일정한 크기의 well을 만들고 선발한 세균을 각각 20 μ l씩 접종하였다. 접종 후 28 °C에서 7일간 배양하였다. Blue color의 배지가 yellow/orange color의 clear zone을 생성하는지 확인하여 siderophore의 생성 여부를 확인하였다.
Hydrogen cyanide: 선발된 세균을 TSA에 glycine (4.4 g/1 l)이 첨가된 배지에 각각 30 μ l씩 도말하였다. 동시에 1% sodium carbonate와 0.5% (w/v) picric acid를 혼합한 용액에 121 °C에서 15분간 멸균한 90-mm filter paper (No. 20)를 침지시킨 후, petri dish 뚜껑에 부착시켜 sealing하였다. 그 후 28 °C에서 72시간 배양하여 뚜껑에 부착된 filter paper가 yellow color에서 red/brown color로 변화하는 것으로 hydrogen cyanide (HCN) 생성 여부를 확인하였다.
Protease production: 선발된 세균의 protease 생성 여부를 확인하기 위하여 실험을 진행하였다. 선발된 세균을 각각 TSB 배지에 28 °C, 130 rpm, 24시간 액체 배양한 후, skim milk agar (skim milk powder 100.0 g, peptone 5.0 g, agar 15.0 g/1 l) 배지에 cork borer (⦶: 5 mm)를 이용하여 일정한 크기의 well을 만들고 배양된 각각의 세균을 20 μ l씩 접종하였다. 28 °C에서 72시간 배양한 후 well 주변의 clear zone을 확인하여, 단백질 분해효소 생성 여부를 확인하였다.
Chitinase production activity: 선발된 세균의 항 진균 물질인 chitinase의 생성 여부를 확인하기 위하여 colloidal chitin을 제조하여 사용하였다. 제조 방법은 chitin from shrimps shells (Sigma-Aldrich, St. Louis, MO, USA) 20 g을 HCl 350 ml에 완전히 녹여 4 °C 냉장고에서 24시간 정치하였다. 그 다음 거즈로 filtering하여 -20 °C에서 보관한 2 l의 ethanol에 빠르게 혼합하였다. Chitin 현탁액을 centrifuge에 10,000 rpm, 20분간 원심분리하였다. 원심분리하여 얻어진 상층액은 버린 후 pellet은 pH가 7.0이 되도록 멸균수로 세척 하였다. 그 후 얻어진 pellet은 동결건조하여 -20 °C 에서 보관하였다. Colloidal chitin agar는 TSA에 colloidal chitin 2% (w/v)를 첨가하여 배지를 제조하였다. Cork borer (⦶: 5 mm) 를 이용하여 well을 만들어 선발된 세균을 각각 20 μ l씩 접종하여 28 °C에서 7일간 배양하였다. Chitinase의 분비는 well 주변부의 clear zone을 측정하여 확인하였다.
결과 및 고찰
수박재배지 및 수박 육묘장에서 샘플 수집 및 미생물 분리. 전국 12지역 94곳의 수박재배지에서 토양 94개, 잎 49개, 꽃 26개, 뿌리 24개 총 193개의 샘플을 수집하였으며, 전국의 25곳의 대형 육묘장에서 123개의 수박 모종 샘플을 수집하였다(Table 1). TSA 배지에서는 수박재배지와 육묘장에서 각각 1,154종과 496종 총 1,650종의 세균이 분리되었으며, YPD 배지에서는 수박재배지와 육묘장에서 각각 799종과 345종 총 1,144종의 세균이 분리되었다(Tables 2, 3). 샘플에서 분리된 총 2,794종의 미생물은 선발한 A. citrulli에 대한 길항성 검정에 사용하였다.
Table 1.
Bacterial isolates from watermelon cultivated at various nurseries of different sites
| Collection site | Soil | Leaf | Flower | Root | Seedling a | Total | |
|---|---|---|---|---|---|---|---|
| Gyeonggi-do | Yangpyeong-gun, Yeoju-si | 12 | 7 | − | 6 | 18 | 43 |
| Gangwon-do | Yanggu-gun, Chuncheon-si, Wonju-si Hongcheon-gun | 10 | 2 | − | − | 21 | 33 |
| Chungcheongbuk-do | Eumseong-gun | 14 | 14 | 5 | 6 | 2 | 41 |
| Chungcheongnam-do | Buyeo-gun, Cheongyang-gun, Gongju-si, Nonsan-si | 17 | 10 | − | 5 | 38 | 70 |
| Jeollabuk-do | Gochang-gun, Jeongeup-si, Wanju-gun | 3 | − | − | − | 8 | 11 |
| Jeollanam-do | Gwangju-si, Naju-si, Hampyeong-gun, Damyang-gun | 4 | 2 | − | 1 | 13 | 20 |
| Gyeongsangbuk-do | Bonghwa-gun, Andong-si, Gumi-si Seongju-gun, Goryeong-gun | 22 | 12 | 11 | 6 | 14 | 65 |
| Gyeongsangnam-do | Geochang-gun, Uiryeong-gun, Changwon-si | 12 | 2 | 10 | − | 9 | 33 |
| Total | 94 | 49 | 26 | 24 | 123 | 316 |
Acidovorax citrulli 분리 및 동정. 수박재배지 및 육묘장 샘플에서 분리된 총 2,794종의 미생물 중 육안 검정과 Aac Immunostrip test 결과 2개의 밴드를 보이는 A. citrulli 후보 균주를 1차 선발하였으며(Fig. 1A), 1차 선발한 균주의 DNA를 추출한 뒤 PCR을 통해 DNA를 증폭하였다. 증폭된 DNA는 전기영동에 사용하였으며, fluorometer를 사용하여 확인한 결과 2개의 균주 모두 360 bp에 밴드를 보이는 것을 확인하였다(Fig. 1B). 분석기관의 capillary electrophoresis sequencing을 통해 동정한 결과 최종적으로 99% 유사성을 가진 2종(HPP-3-3B, HPP-9-4B)의 A. citrulli를 선발하였다(Fig. 1C, D). 선발된 HPP-3-3B과 HPP-9-4B 의 GenBank accession 번호는 각각 MW788378과 MW788377이다.
Fig. 1.
Two bacterial fruit blotch (BFB) isolates selected among the microorganisms collected from various sites. Collected BFB isolates were ana-lyzed by immunostrip test followed by phylogenetic relationship. (A) Two selected bacterial isolates showing double band in the immunostrip test. (B) Polymerase chain reaction test result showing a band at 360 bp. (C) Phylogenetic tree of isolate HPP21-3-3B showing the similarity level of 99% with Acidovorax citrulli. (D) Phylogenetic tree of the study isolate HPP21-9-4 showing the similarity level of 99% with A. citrulli.

박과 작물에 과실썩음병을 유발하는 A. citrulli는 과실에 발생이 많으며, 육묘기부터 발생해 접목 후 확산하는 것으로 알려져 있으며(Hopkins와 Thompson, 2002), 그 외에도 수분, 관수, 정식 후 적심 등의 재배관리 작업을 할 때 전염의 우려가 있다(Noh와 Choi, 2015). 본 연구에서 분리한 BFB (HPP-3-3B, HPP-9-4B)는 모두 육묘장에서 채취한 식물체에서 분리된 미생물이었으며(Table 2), 파종, 접목 등의 작업 과정에서 BFB가 식물체에 감염된 것으로 판단된다.
Table 2.
Number of bacteria isolated from each sample on TSA media
| Collection site | Total no. of isolated bacteria from each sample | ||||||
|---|---|---|---|---|---|---|---|
| Soil | Leaf | Flower | Root | Seedling a | Total | ||
| Gyeonggi-do | Yangpyeong-gun, Yeoju-si | 67 | 22 | − | 24 | 53 | 166 |
| Gangwon-do | Yanggu-gun, Chuncheon-si, Wonju-si Hongcheon-gun | 95 | 8 | − | − | 90 | 193 |
| Chungcheongbuk-do | Eumseong-gun | 151 | 132 | 7 | 46 | 4 | 340 |
| Chungcheongnam-do | Buyeo-gun, Cheongyang-gun, Gongju-si, Nonsan-si | 84 | 62 | − | 24 | 162 | 332 |
| Jeollabuk-do | Gochang-gun, Jeongeup-si, Wanju-gun | 20 | 6 | − | 4 | 31 | 61 |
| Jeollanam-do | Gwangju-si, Naju-si, Hampyeong-gun, Damyang-gun | 23 | − | − | − | 57 (2) b | 80 |
| Gyeongsangbuk-do | Bonghwa-gun, Andong-si, Gumi-si Seongju-gun, Goryeong-gun | 158 | 36 | 28 | 16 | 68 | 306 |
| Gyeongsangnam-do | Geochang-gun, Uiryeong-gun, Changwon-si | 100 | 9 | 32 | − | 31 | 172 |
| Total | 698 | 275 | 67 | 114 | 496 | 1,650 | |
Table 3.
Number of bacteria isolated from each sample on YPD media
| Collection site | Total no. of isolated bacteria from each sample | ||||||
|---|---|---|---|---|---|---|---|
| Soil | Leaf | Flower | Root | Seedling a | Total | ||
| Gyeonggi-do | Yangpyeong-gun, Yeoju-si | 70 | 25 | − | 22 | 37 | 154 |
| Gangwon-do | Yanggu-gun, Chuncheon-si, Wonju-si Hongcheon-gun | 72 | 10 | − | − | 23 | 105 |
| Chungcheongbuk-do | Eumseong-gun | 51 | − | − | − | 26 | 77 |
| Chungcheongnam-do | Buyeo-gun, Cheongyang-gun, Gongju-si, Nonsan-si | 64 | 42 | − | 22 | 69 | 197 |
| Jeollabuk-do | Gochang-gun, Jeongeup-si, Wanju-gun | 13 | 2 | − | 2 | 108 | 125 |
| Jeollanam-do | Gwangju-si, Naju-si, Hampyeong-gun, Damyang-gun | 12 | 2 | − | 2 | 1 | 17 |
| Gyeongsangbuk-do | Bonghwa-gun, Andong-si, Gumi-si Seongju-gun, Goryeong-gun | 172 | 24 | 28 | 21 | 11 | 256 |
| Gyeongsangnam-do | Geochang-gun, Uiryeong-gun, Changwon-si | 103 | 9 | 31 | − | 70 | 213 |
| Total | 557 | 114 | 59 | 69 | 345 | 1,144 | |
분리된 A. citrulli에 대한 길항 미생물 선발. 샘플에서 분리된 2,792종의 미생물과 선발한 BFB (A. citrulli)에 대한 1차 길항성 검정 결과 38종의 길항 후보 균주가 선발되었다. 1차 선발한 길항 미생물에 대한 2차 길항성 검정 결과 HPP-3-3B에 길항을 가지는 미생물은 24종이었으며, HPP-9-4B는 13종의 미생물이 길항을 가지는 것으로 확인되어, 총 28종의 길항 세균을 선발하였다(Figs. 2, 3). 선발한 길항 세균 중 BNPL-6-3B, HYGPL-1-3B, TIPL-6-1B, YGMP-2-7Y는 2종의 BFB균 모두에서 강한 길항성을 보였다. 선발한 길항 세균은 HPP-9-4B보다 HPP-3-3B에서 더 강한 길항성을 보였으며, Bacillus velezensis로 확인된 NJSP-5-1Y, JSP-1-1Y는 HPP-3-3B에 대해 강한 길항성을 나타냈지만, HPP-9-4B에서는 길항성을 나타내지 않았다(Table 4). 선발한 28종의 길항 세균의 동정 결과 Bacillus 16종, Peudomonas 7종, Pantoea, Arthrobacter, Lelliottia, Beijerinckia, Curtobacterium이 각각 1종씩 동정되었다(Table 4).
Fig. 2.
Antagonistic effect of selected bacterial isolates against HPP-3-3B. C, control; 1, CB20L-4-5B; 2, CB20L-9-1B; 3, CB20L-11-5B; 4, CB20L-12-2B; 5, BNPL-3-3B; 6, BNPL-6-3B; 7, HYGPL-1-3B; 8, TIPL-1-5B; 9, TIPL-4-2B; 10, TIPL-6-1B; 11, TIPL-7-4B; 12, TIPL-8-1B; 13, KB20F-2-1Y; 14, KB20L-7-2Y; 15, KB20L-11-1Y; 16, KW20S-3-2Y; 17, JSP-1-1Y; 18, JSP-4-2Y; 19, KSNHP-1-3Y; 20, NJSP-1-5Y; 21, NJSP-1Y; 22, YGP-1-3Y; 23, YGP-2-5Y; 24, YGWMP-2-7Y.

Fig. 3.
Antagonistic effect of selected bacterial isolates against HPP-9-4B. C, control; 1, CB20L-9-1B; 2, CB20L-11-5B; 3, CB20R-2-5B; 4, BNPL-6-3B; 5, HYGP-1-3B; 6, TIPL-1-5B; 7, TIPL-4-2B; 8, TIPL-6-1B; 9, TIPL-8-1B; 10, KB20F-2-1Y; 11, KB20L-3-2Y; 12, KB20L-7-2Y; 13, KB20L-11-1Y; 14, YGWMP-2-7Y.
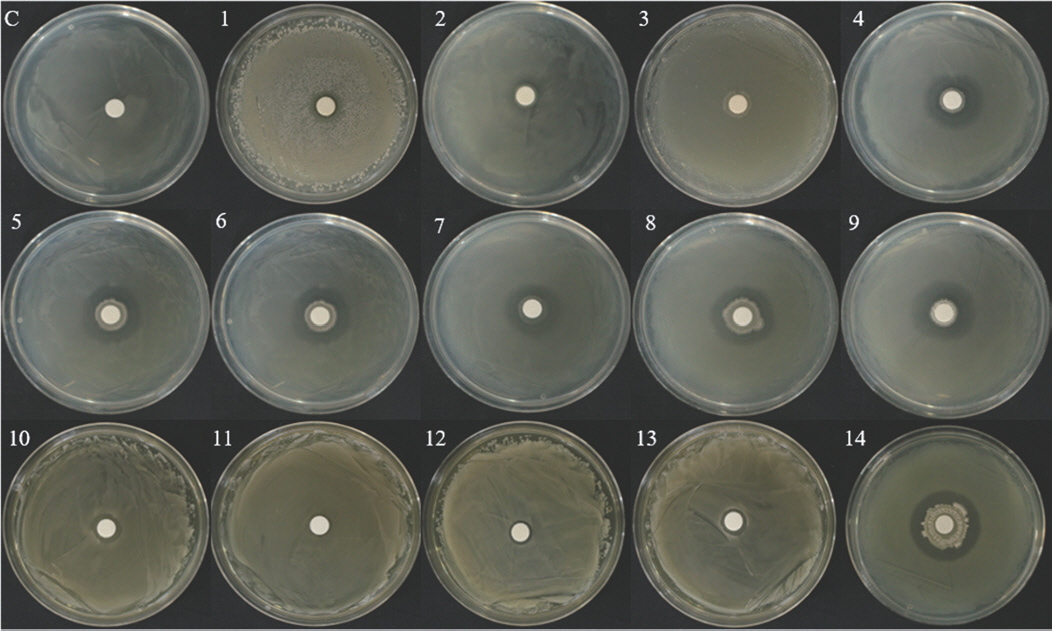
Table 4.
In vitro antagonistic tests against bacterial fruit blotch with selected bacterial isolates from each sample
최근 B. velezensis와 Peudomonas brassicacearum 균주를 사용하여 토마토 시들음병을 유발하는 R. solanacearum을 효과적으로 억제한다고 보고하였으며(Elsayed 등, 2020), Ahmad 등(2019)은 Bacillus subtilis 균주를 발효 기술을 사용하여 생물 방제제를 생산하여 Rhizoctonia solani에 의한 땅콩의 뿌리 썩음을 억제하였다고 보고하였다. Pantoea는 근권 토양에서 식물성장 촉진물질인 항진균물질과 auxin을 동시에 생산한다고 보고되었다(Kwon 등 2004). 또한, Jung 등(2007)은 Pantoea agglomerans는 Fusarium oxysporum과 Colletotrichum gloeospo-rioides의 생육을 억제하였으며, Bacillus megaterium은 Botrytis cinerea, Sclerotinia sclerotiorum 및 Phythium ultimum의 생육을 억제한다고 보고하였다.
선발된 길항 미생물의 생화학적 검증.
Ammonia production: 선발된 세균의 ammonia 생성 여부를 확인하기 위하여 실험을 진행하였다. 수박재배지에서 선발한 11종의 세균 모두 ammonia를 생성하는 것으로 나타났으며(Table 5, Fig. 4), 육묘장에서 선발한 17종의 세균 중 JSP-1-1Y를 제외한 16종 모두 ammonia를 생성하는 것으로 확인되었다.
Fig. 4.
Ammonia production in 28 antagonistic bacteria. The color variation was confirmed in 1 ml Nessler's reagent. C, control; 1, CB20R-2-5B; 2, CB20L-11-5B; 3, CB20L-9-1B; 4, CB20L-12-2B; 5, CB20L-4-5B; 6, KB20L-7-2B; 7, KB20L-3-2Y; 8, KB20L-7-2Y; 9, KB20L-11-1Y; 10, KB20F-2-1Y; 11, KW20S-3-2Y; 12, BNPL-3-3B; 13, BNPL-6-3B; 14, TIPL-1-5B; 15, TIPL-4-2B; 16, TIPL-6-1B; 17, TIPL-7-4B; 18, TIPL-8-1B; 19, HYGP-1-3B; 20, JIP-5-2B; 21, NJSP-1-5Y; 22, NJSP-5-1Y; 23, YGMP-2-7Y; 24, KSNHP-1-3Y; 25, YGP-1-3Y; 26, YGP-2-5Y; 27, YGP-4-2Y.

Table 5.
PGPR traits and antagonistic traits of selected 28 antagonistic bacterial isolates
질소는 식물생육에 있어서 필요한 원소 중 하나로, 식물의 신장, 엽면적 확장 및 분열 촉진에 영향을 주는 것으로 알려져 있어 식물생육에 가장 중요한 필수 원소 중 하나이다(Jang 등, 2017). Ahmad 등(2008)에 따르면, 암모니아의 생산은 식물의 성장의 간접적인 촉진 특성 중 하나로 간주된다. 질산환원효소는 질소고정세균에서 NH3에 고정된 N2를 환원시키는 데 중요한 역할을 하며, 고정된 암모니아는 glutamate와 glutamine으로 동화되어 세포에서 아미노산, 핵산 및 단백질의 형성에 중요한 역할을 한다. 박테리아 및 용해 후 이러한 화합물은 환경으로 방출되어 식물 등의 유기체에 의해 질소 공급원으로 사용된다(Hartono 등, 2016). 암모니아를 생산하는 미생물로 토마토 식물에 접종하였을 경우 줄기 길이, 잎 수, 잎 면적 등의 명확한 성장 개선을 보였다(Essalimi 등, 2022).
Phosphate solubilization: Cappuccino와 Sherman (1992)의 실험 방법을 참고하여, 선발된 세균의 불용성 인산 가용화 능력을 확인하는 실험을 진행하였으며, 실험 결과 CB20R-2-5B, CB20L-11-5B, KB20F-2-1Y, KW20S-3-2Y, BNPL-3-3B, BNPL-6-3B, TIPL-4-2B, TIPL-6-1B, TIPL-7-4B의 9개 균주가 clear zone을 형성하는 것으로 나타났으며, 각각 0.98 cm, 1.61 cm, 1.07 cm, 1.99 cm, 0.31 cm, 0.78 cm, 1.15 cm, 1.10 cm, 0.99 cm의 인산 가용화 능력을 확인하였다(Table 5, Fig. 5).
Fig. 5.
Solubilization of inorganic phosphate. The formation of clear zones around the bacterial inoculation point were tested on Pikovskaya agar. C, control; 1, CB20R-2-5B; 2, CB20L-11-5B; 3, KB20F-2-1Y; 4, KW20S-3-2Y; 5, BNPL-3-3B; 6, BNPL-6-3B; 7, TIPL-4-2B; 8, TIPL-6-1B; 9, TIPL-7-4B.

Phosphate는 식물 성장에 필요한 영양소 중에서 질소 다음으로 중요한 영양요소이며 식물의 거의 모든 주요 대사 과정에서 중요한 역할을 담당하고 있다(Khan 등, 2010). 인산 분해 미생물은 식물의 대표적인 생장 호르몬인 지베렐린과 인돌아세트산을 생성하여 식물 생장을 촉진한다고 보고되었다(Yun과 Cheong, 2016). Xue 등(2019)은 인산 가용화 미생물이 유기물을 합성하고 식물 바이오매스를 증가시키기 위한 영양소의 효율적인 흡수 및 사용을 촉진하여 식물 성장을 촉진하고 뿌리줄기와 잎 바이오매스를 증가시킬 수 있음을 입증하였다. 또한, 유용 미생물을 토양에 접종하여 유용 미생물의 인산 분해 능력이 땅콩의 성장 촉진에 영향을 주는 것을 입증하였다(Wang 등, 2021).
Starch hydrolysis: 전분 가수분해 효소를 생성하는지 확인하기 위하여 starch agar에 선발한 세균을 접종하여 28 °C에서 48시간 배양한 후 iodine을 부어 전분이 가수분해되어 흑자색으로 염색되지 않은 clear zone을 확인하였다. 수박재배지에서 선발한 11종의 세균 모두 전분 가수분해 효소를 생성하지 않았으나, 육묘장에서 선발한 17종의 세균은 모두 전분 가수분해 효소를 생성하였다(Table 5, Fig. 6).
Fig. 6.
Starch hydrolysis by 28 antagonistic bacteria. The clear zone formation was monitored on the starch agar media. C, control; 1, BNPL-3-3B; 2, BNPL-6-3B; 3, TIPL-1-5B; 4, TIPL-4-2B; 5, TIPL-6-1B; 6, TIPL-7-4B; 7, TIPL-8-1B; 8, HYGP-1-3B; 9, JIP-5-2B; 10, NJSP-1-5Y; 11, NJSP-5-1Y; 12, JSP-1-1Y; 13, YGMP-2-7Y; 14, KSNHP-1-3Y; 15, YGP-1-3Y; 16, YGP-2-5Y; 17, YGP-4-2Y.
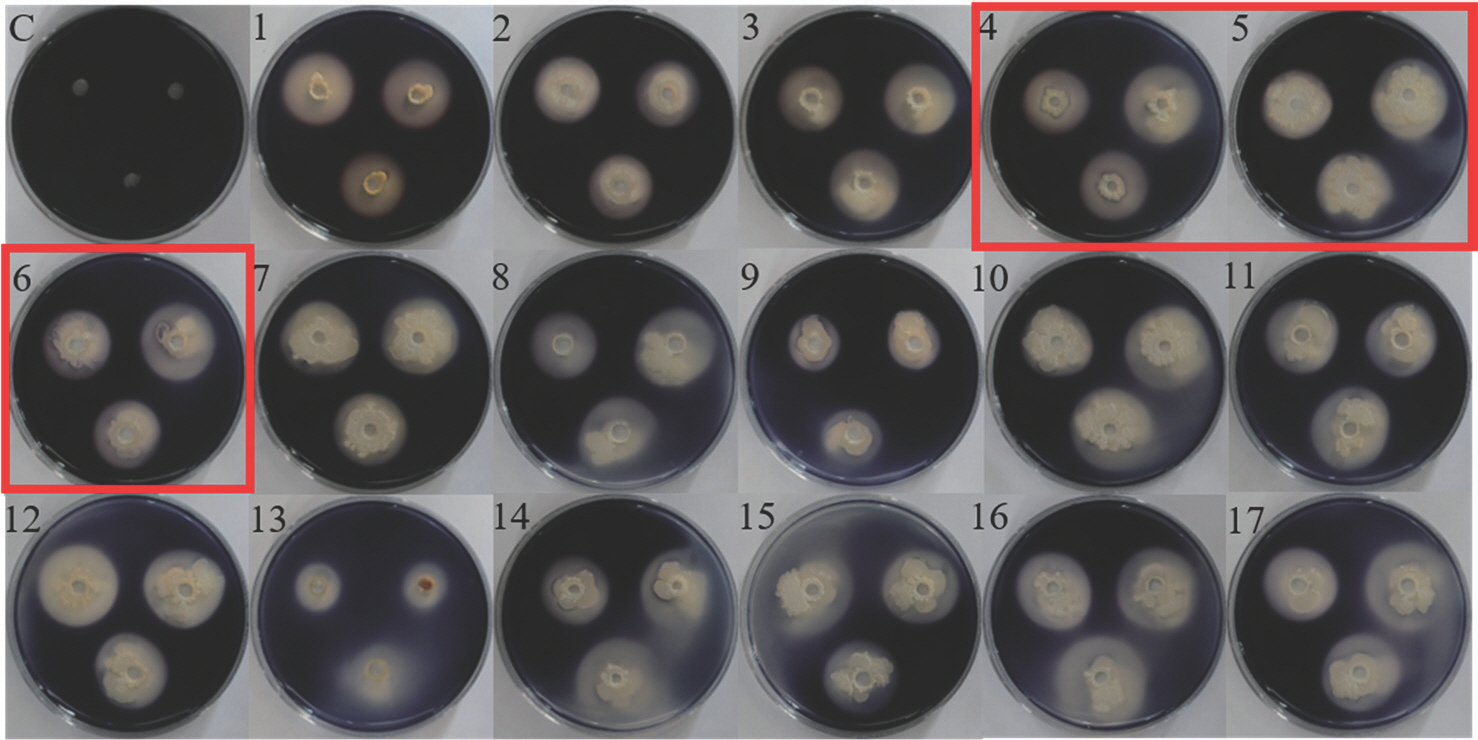
전분은 식물의 생장 과정에서 중요한 역할을 하며, 에너지 대사를 위해 잎의 엽록체에서 합성되고 성장 단계를 유지하기 위해 줄기, 뿌리, 종자 등의 비광합성기관에서도 합성된다(Vajravijayan 등, 2018). 전분 가수분해 효소의 생산은 박테리아가 다른 미생물의 성장을 억제하는 데 사용하는 일반적인 메커니즘으로 알려져 있다. 연구에 따르면, 생강에서 분리한 전분 가수분해 효소를 생성하는 PGPR Bacillus amyloliquefaciens를 생강에 처리하여 발아를 향상시키고 무름병 발병을 억제하였다(Dinesh 등, 2015).
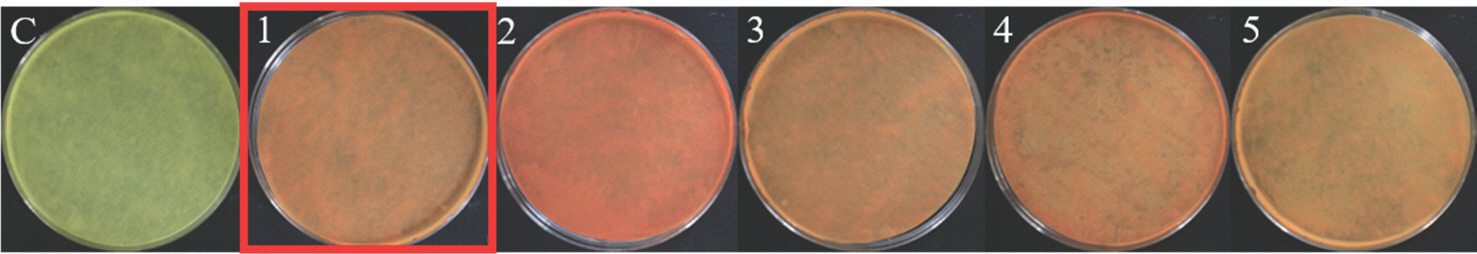
Siderophore production: 선발된 세균이 siderophore를 생성하는지 확인하기 위하여 CAS가 함유된 CAS blue agar에 균주들을 접종하여 28 °C에서 7일간 배양하여 clear zone을 확인하였다. 확인 결과 선발된 균주 중 23개의 세균이 yellow/orange color의 clear zone을 형성하였고, BNPL-6-3B가 2.40-cm clear zone을 형성하여 23개의 균주 중 가장 높은 siderophore 생성능을 확인하였다(Table 5, Fig. 7).
Fig. 7.
Siderophore production. The orange/yellow color around the bacteria inoculation point on Chrome Azurol S agar media indicates siderophore formation. C, control; 1, CB20R-2-5B; 2, CB20L-11-5B; 3, CB20L-9-1B; 4, CB20L-4-5B; 5, KB20L-7-2B; 6, KB20L-7-2Y; 7, KB20L-11-1Y; 8, KB20F-2-1Y; 9, BNPL-3-3B; 10, BNPL-6-3B; 11, TIPL-1-5B; 12, TIPL-4-2B; 13, TIPL-6-1B; 14, TIPL-7-4B; 15, TIPL-8-1B; 16, NJSP-1-5Y; 17, NJSP-5-1Y; 18, JSP-1-1Y; 19, KSNHP-1-3Y; 20, YGP-1-3Y; 21, YGP-2-5Y; 22, YGP-4-2Y.

Siderophore는 미생물이 철을 흡수하기 위해 체외로 분비하는 펩티드성 물질로서 식물 생장 촉진과 상관관계가 있다는 여러 연구결과가 있으며, siderophore의 생성은 PGPR의 핵심 요소 중 하나이다(Crowley 등, 1991). Siderophore는 킬레이트화 반응을 통해 중금속에 의해 유발되는 철 결핍을 감소시켜 식물이 헴 및 엽록소와 같은 광합성 화합물을 합성하는 데 도움을 주는 것으로 알려져 있다(Ferreira 등, 2019). Kumar와 Dube (1992)는 완두콩과 대두 종자에 siderophore를 생산하는 Pseudomonas를 접종하면 발아, 성장 및 수확량이 증가한다고 보고했다. 또한, Pseudomonas에 의해 생성된 siderophores 는 Fusarium oxysporum, Fusarium udum 및 Aspergillus niger에 의해 유발되는 질병을 제어하고 싹과 뿌리의 성장을 자극한다고 보고했다(Trapet 등, 2016).
HCN production: HCN을 형성하는지 확인하기 위하여 fiter paper 색의 변화를 관찰하였다. 실험 결과 CB20R-2-5B, CB20L-4-5B, KB20L-7-2B, KB20L-3-2Y, KB20L-11-1Y 5개의 균주가 yellow color에서 red/brown color로 변한 것을 확인하였으며, 이것으로 5개의 균주가 HCN을 생성하는 것을 확인하였다(Table 5, Fig. 8).
Fig. 8.
Hydrogen cyanide production by 28 antagonistic bacteria. The filter paper assay with 0.5% picric acid and 1% NaCl was employed to demonstrate HCN production. C, control; 1, CB20R-2-5B; 2, CB20L-4-5B; 3, KB20L-7-2B; 4, KB20L-3-2Y; 5, KB20L-11-1Y.
HCN을 생성하는 세균은 식물병원성 세균을 억제하는 것으로 알려져 있으며(Voisard 등, 1989), 곰팡이 억제에 효과적이며, 곰팡이 호흡 사슬에서 cytochrome-C 말단 산화효소를 억제하고 곰팡이 대사에 극심한 손상을 주는 것으로 알려져 있다(Singh 등, 2019). 특히, Jung 등(2012)은 선충류에 의한 작물 피해가 많은 토양에서 HCN 농축을 사용하면 좋은 효과를 얻을 것으로 기대된다고 하였다. Abd El-Rahman 등(2019)은 HCN 을 생성하는 PGPR Pseudomonas와 Bacillus를 Meloidogyne incognita에 감염된 토양에 접종하여 토마토의 성장 매개변수를 증가시켰다.
Protease production: Protease (단백질분해효소) 생산 능력을 확인하기 위하여 실험을 진행한 결과 수박재배지와 육묘장에서 선발한 28개의 균주 중 총 23개의 균주가 protease를 생산하는 것으로 확인되었다. 23개 균주는 평균 1.97 cm 크기의 clear zone을 형성하며 protease를 생산하는 것을 확인하였다. 균주 중 KB20L-7-2B가 2.90 cm의 clear zone을 형성하여 가장 많은 protease를 생산하는 것으로 나타났고, CB20L-4-5B가 1.59 cm의 clear zone을 형성하여 가장 적은 단백질 분해효소를 생산하는 것으로 나타났다(Table 5, Fig. 9).
Fig. 9.
Protein hydrolysis by 28 antagonistic bacteria. The clear zone on skim milk agar media represented protein hydrolysis. C, control; 1, CB20R-2-5B; 2, CB20L-11-5B; 3, CB20L-4-5B; 4, KB20L-7-2B; 5, KB20L-7-2Y; 6, KB20L-11-1Y; 7, KB20F-2-1Y; 8, KW20S-3-2Y; 9, BNPL-3-3B; 10, BNPL-6-3B; 11, TIPL-1-5B; 12, TIPL-4-2B; 13, TIPL-6-1B; 14, TIPL-7-4B; 15, TIPL-8-1B; 16, HYGP-1-3B; 17, NJSP-1-5Y; 18, NJSP-5-1Y; 19, JSP-1-1Y; 20, KSNHP-1-3Y; 21, YGP-1-3Y; 22, YGP-2-5Y; 23, YGP-4-2Y.

Protease는 단백질을 이루고 있는 아미노산간의 펩티드 결합을 가수분해하는 효소이다. Protease는 식물의 발달조절이나 광합성에 영향을 주는 것으로 알려져 있으며(Clemente 등, 2019), bacteria와 fungi의 단백질의 경우 생태계의 탄소와 질소의 순환에 매우 중요한 역할을 하는 것으로 알려져 있다(Fricke 등, 2001). 특히, 식물에서 해충과 병원성균에 대한 방어제 및 억제제로 사용하기 위해 이용되고 있다고 보고하였다(Clemente 등, 2019).
Chitinase production: 선발된 세균이 chitinase 효소를 생성하는지 확인하기 위하여 실험을 진행하였다. 본 연구에서 선발한 28종의 길항 미생물은 실험 결과 chitinase 효소를 생성하지 않았다(Table 5). Chitinase는 선충의 발생을 억제하며, fungi 세포벽의 주성분인 chitin의 가수분해를 촉매하여 병원성 곰팡이의 생장을 억제하는 것으로 알려져 있다(Mahadevan과 Crawford, 1997; Punja와 Zhang, 1993). 최근에는 chitinase 효소를 이용하여 병해충관리나 토양개량 및 작물생육에 도움이 되는 친환경 유기농 자재를 개발하고 있다(Jung, 2020).
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






