



서 론
트리트케일은 밀을 모본으로 하고, 호밀(Secale cereal)을 부본하여 얻은 잡종의 염색체를 배가 시킨 복이질배수체 작물로써 밀의 양질성과 호밀의 우수한 생장능력 및 내재저항성을 결합한 작물이다. 가축이 좋아하는 밀의 품질과 추위에 잘 견디는 호밀의 특성을 지니고 있는 대표적인 겨울철 사료작물이다. 트리티케일은 듀럼밀(Triticum turgidum)과 호밀을 교잡한 6배체와 빵밀(Triticum aestivum)과 호밀을 교잡한 8배체가 있으나 생육과 종자 생산성에 있어 안정성이 우수한 6배체가 널리 재배되고 있다(Hills 등, 2007; Mergoum 등, 2009; Simmonds, 1976). 트리티케일은 독일, 프랑스, 폴란드 등에서는 팬케이크와 같은 식용으로 사용할 뿐만 아니라 청예, 건초, 사일리지 형태의 가축사료로 사용되고 있다(Ciba, 1983; Mergoum 등, 2009). 최근에는 에탄올 추출용으로 이용되고 있기도 하며, 국내에서는 축산농가의 조사료 생산에 이용할 목적으로 도입되었다(Heo 등, 2002). 트리티케일은 식물체의 잎 비율이 높고, 이탈리안라이그라스, 호밀, 청보리 등과 비교해 이삭이 팰 때의 조단백질 함량이 더 높고, 가소화양분의 함량도 더 많아 가축 기호성이 높은 장점이 있으며, 이상기후에 민감한 다른 월동 작물에 비해 생산성이 높고 안정성이 우수하여 사료 작물로서 그 수요가 증가하고 있는 추세이다. 국내에서도 신기호밀(Youn 등, 1986), 신영(Heo 등, 2002), 조성(Han 등, 2012), 신성(Han 등, 2016), 세영(Han 등, 2017), 그리고 조영(Han 등, 2019) 등이 개발되어 보급하고 있다.
점차 식품 및 조사료로써의 중요성이 증가하고 있는 트리티케일에서도 밀, 보리, 귀리 등과 같이 붉은곰팡이에 의한 감염 피해가 발생하고 있다(Arseniuk 등, 1993). 맥류 붉은곰팡이병은 아시아, 북미, 유럽 등 전 세계적으로 밀, 보리, 옥수수, 쌀, 귀리 등의 다양한 곡류에 발병하여 많은 피해를 주는 난방제병이다. 붉은곰팡이병의 감염은 수량 감소와 품질 저하의 원인이 되어 경제적 손실의 초래할뿐만 아니라 붉은곰팡이균이 생산하는 곰팡이 독소로 인해 인축에 구토, 설사, 면역력 저하, 암 등 다양한 질병의 원인이 되기도 한다. 붉은곰팡이균이 생성하는 주요 독소로는 deoxynivalenol (DON), nivalenol (NIV), zearalenone 등이 보고되어 있다(Desjardins, 2006; Leslie와 Summerell, 2006). 전세계적으로 맥류 붉은곰팡이병을 일으키는 다양한 균주들이 알려져있는데, 그 중 주요 원인균은 Fusarium graminearum, F. pseudograminearum, F. culmorum, F. avenaceum 등의 균주들이 보고되었고, 이들이 복합체를 이루어 존재하는 것으로 알려져 있다. 이런 균주들은 지역별로 종의 분포가 다르며, 동일한 종이라 하더라도 계통발생학적으로 다양한 것으로 알려져 있다(O’Donnell 등, 2004; Yang 등, 2008). 전 세계적으로 발생하는 붉은곰팡이병은 국내에서도 밀, 보리, 옥수수, 쌀, 귀리에서 병이 발생 하는 것으로 알려져 있다(Choi 등, 2019; Kim 등, 2016; Lee 등, 2010; Ryu 등, 2011). 국내에서 밀, 보리, 쌀, 귀리 등에서 발생하는 붉은곰팡이병에 대한 연구는 많이 보고되었으나, 트리티케일에서 발생하는 붉은곰팡이병에 대한 연구는 미흡한 실정이다. 본 연구에서는 국내에서 육성 및 재배되고 있는 주요 트리티케일 품종에 발생하는 붉은곰팡이병의 주요 원인균과 독소의 다양성 분석하고, 병 방제를 위해 사용 가능한 약제들의 약제 내성 발생 유무를 구명하고자 하였다.
재료 및 방법
병 발생률 조사 및 균주 분리. 2021년 국립식량과학원 중부 작물부(54, Seoho-ro, Gwonseon-gu, Suwon-si)의 트리티케일 재배 포장에서 5개 품종의 트리티케일의 붉은곰팡이병 발생률 조사와 병징이 보이는 종실을 수집하였다. 붉은곰팡이병 발생률 조사는 농촌진흥청 과학기술연구조사분석기준에 의거하여 진행하였다. 트리티케일의 품종별로 재배되는 포장에서 각 5개 지점을 무작위로 지름 30 cm 원형틀을 이용하여 틀 안에 트리티케일 총 수량 대비 병 발생 개체 수를 측정하였다. 또한, 병 발생 개체의 각각의 종실에서 병 발생 낱알을 측정하여 종실 전체 낱알 대비 병 발생 낱알률을 측정하였다. 수집한 병 발생 개체 시료로부터 병원균을 분리하기 위해 1% NaOCl 용액으로 3분간 표면 살균 후 멸균수로 2회 세척을 하였다. 이후, clean bench 안에서 멸균된 여과지가 들어있는 plate 위에 치상한 후 5-7일 동안 습실 처리하였다. 이 중 Fusarium spp.으로 예상되는 균의 단포자를 취하여 potato dextrose agar (PDA; rifampicin 100 μg/ml)에서 배양하였다.
균 동정 및 독소 화학형 분석. Fusarium spp. 균주를 확인하기 위해 포자 형성이 용이한 카네이션잎 한천 배지에 균총을 접종 후 24시간의 광조건에서 7‒10일간 배양한 후 near ultraviolet이 12 hr/12 hr 조사되는 25oC 항온기에서 24시간 배양하였다. 배지에서 형성된 소형 및 대형분생포자의 유무 등 형태적 특성과 PDA 배지에서 형성된 균총의 색 등으로 균주의 배양적 특성을 조사하였다. 그 결과를 Fusarium 분류 메뉴얼(Leslie와 Summerell, 2006)에 따라 1차적으로 종(species) 수준으로 구분하였다. 이후 균의 분자적 동정을 위해 internal transcribed spacer 1 (ITS1) (White 등, 1990), translation elongation factor 1α (TEF-1α) (O′Donnell 등, 1998) 유전자의 염기서열을 분석하였다. 유전자 염기서열은 MEGA-X프로그램의 UPGMA (unweighted pair group method with arithmetic mean)법으로 계통수를 작성하였다. 계통수 분석에 사용된 F. asiatium NRRL 26156등의 ITS1, TEF-1α 유전자의 염기서열은 National Center for Biotechnology Information (NCBI)에 등록된 유전정보를 이용하여 분석하였다. Trichothecene은 TRI7과 TRI13 유전자(Ward 등, 2008)을 이용하여 Fusarium spp. 균주의 독소화학형 분석을 진행하였다.
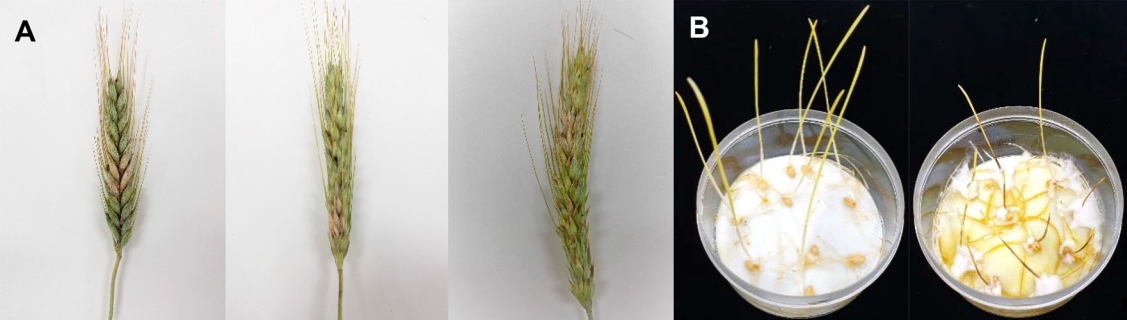
병원성검정. 붉은곰팡이병 증상을 나타낸 트리티케일에서 분리한 균주들을 대상으로 트리티케일에 대한 병원성 검정을 진행하였다. 트르티케일에서 분리된 72개의 균주를 다시 트리티케일(신영)유묘에 접종하였으며, 대조구로 이미 기존 밀에서 병원성 검정이 완료된 F. graminearum PH-1과 F. asiaticum 18DG11를 대조구로 이용하여 접종실험을 실시하였다. 트리티케일에서 병원성 검정이 완료된 균주 중 7균주를 선발하여 트리티케일(신영), 밀(조경)에 대하여 병원성 비교 검정을 진행하였다. 병 검정을 위해 사용된 균주는 carnation leaf agar 배지에서 7‒10일 동안 25oC 항온기에서 배양 후 스크래퍼를 이용하여 균사 표면을 긁어낸 후 NUV 선반에서 24시간 배양하였다. 형성된 포자는 수거하여 1×105/ml 포자 농도로 접종원을 만들었다. 유묘기에 접종을 위해 트리티케일 및 밀 종자를 NaOCl 용액에 5분 동안 종자 소독을 진행 후 멸균수로 3회에 걸쳐 씻어내었다. 살균이 완료된 종자를 멸균된 여과지가 놓인 플레이트에 치상하여 3일동안 암실에서 발아를 진행하였다. 이후 4일 동안 생육상에서 명과 암 12 hr/12 hr 간격으로 배양을 진행하였다. 이후, F. asiaticum 18DG11을 포함한 균주들의 접종원 5 ml를 유묘가 자란 플레이트에 접종하였다. 이후 7일간 25oC 생육 상에서 배양 후 병 심각도를 측정하였다. 병의 심각도는 병의 발생 정도에 따라 0에서 5단계로 측정하였다(0단계: 뿌리 및 줄기 등에 병징이 나타나지 않음, 1단계: 뿌리와 줄기 밑 부분에 아주 작게 병징이 나타남, 2단계: 병징이 줄기 밑부분에 눈에 띄게 나타남, 3단계: 병징이 줄기의 중간까지 발생함, 4단계: 병징이 줄기 2/3이상 발생함, 5단계: 병이 심하게 발생하여 뿌리 줄기 등이 고사됨).
농약저항성 발생 확인. 밀과 보리에 사용 가능하도록 등록되어 있는 농약인 captan, fludioxonil, difenoconazole· propiconazole 및 hexaconazole을 농약저항성 실험에 사용하였다. 트리티케일로부터 분리된 72개의 Fusarium spp.의 균주를 PDA 배지에 배양 후 직경 5 mm 코르크보러로 취하여 권장 농도의 농약이 들어간 배지에 올려놓고 7-10일 동안 25oC 항온기에서 배양하였다. 농약을 넣지 않은 균주와 크기를 비교하여 균주별 농약에 대한 저항성 발생 여부를 확인하였다. 사용한 농약의 농도는 각 농약의 권장 농도로 사용하였다. Captan의 경우 2,000 ppm, hexaconazole 500 ppm, fludioxonil은 1,000 ppm을 사용했으며, difenoconazole·propiconazole 합제는 1,000 ppm 농도로 사용하였다.
결 과
트리티케일 품종별 붉은곰팡이병 발생 현황. 국립식량과학원 중부작물부 포장에서 재배되고 있는 트리티케일 조성, 신영, 신조성, 신성, 세영 품종의 붉은곰팡이병 발생 상황을 조사하였다(Fig. 1). 병 발생 조사 결과 병 발생 이삭률은 각각 7.21%, 14.53%, 8.53%, 10.05%, 그리고 5.60%로 나타났으며, 35.99%, 30.3%, 37.22%, 14.83%, 그리고 26.78%의 병 발생 낱알률을 나타내었다. 전체 트리티케일 재배포장의 전체 환산 병 발생률은 평균 3.52%로 나타났다(Table 1).
붉은곰팡이 병 발생 원인균 동정 및 다양성 분석. 트리티케일 5개 품종의 붉은곰팡이병이 발생한 개체로부터 곰팡이 균주를 분리한 결과 형태학적으로 3가지의 곰팡이가 분리되었다(Table 2). 분자동정결과 Penicillium spp. 6.59%, Aspergillus spp. 14.28% 그리고 Fusarium spp. 균주가 79.12%로 분리되어 Fusarium spp. 균주가 주요 병원균으로 분리 되었다(Table 2). 분리된 Fusarium spp. 균주 중 병원성 검정을 통해 다양한 병원성 반응을 보인 15개 균주를 선발하여 ITS1 영역과 TEF-1α 유전자의 염기서열을 분석하였다. 유전자의 염기서열 분석 결과, 15개의 균주 모두 F. asiaticum clade에 속하는 것으로 분류되었다(Fig. 2). 트리티케일에서 분리, 동정된 F. asiaticum은 트리티케일뿐만 아니라 쌀, 보리, 밀, 귀리 등에서도 우점종으로 분리되었다(Choi 등, 2019; Lee 등, 2010). 옥수수의 경우 지역에 따라 붉은곰팡이병의 우점균이 달리 나타났는데 강원도 지역의 경우 DON 생성 F. graminearum이 우점종이였으나, 충북의 경우 NIV 생성 F. asiaticum이 우점종이였다(Lee 등, 2010). 쌀의 경우 F. asiaticum, F. graminearum 외에 F. culmorum, Fusarium vorosii 등이 붉은곰팡이병의 원인균으로 분리되었으며, F. vorosii는 옥수수, 보리에서도 원인균으로 분리 되었다(Kim 등, 2016; Lee 등, 2016). 붉은곰팡이병에는 다양한 원인균이 존재하지만 국내에서 재배되는 쌀과 잡곡 등에서 발생하는 붉은곰팡이병의 주요 원인균은 F. asiaticum으로 판단된다. 본 실험에서는 병 조사 및 균 분리 포장이 한 곳으로 한정되어 지역별 우점균이라고 판단하기 어려우나 F. asiaticum이 트리티케일의 붉은곰팡이병의 원인균임을 확인한 결과이다.
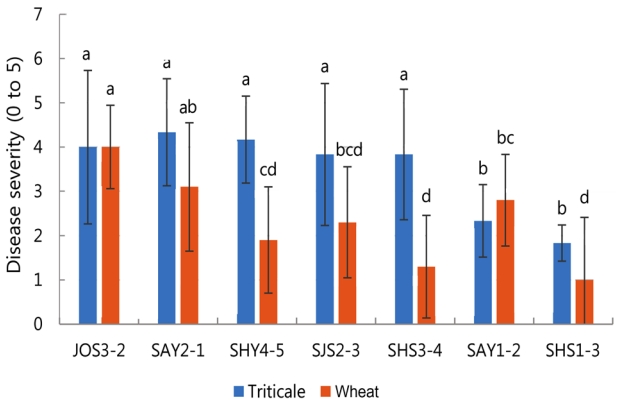
병원성 검정 결과. 트리티케일에서 분리한 균주의 병원성 검정을 진행하였다. 트리티케일에서 분리 및 동정된 균주는 총 72개로 기존 밀과 보리에서 병원성 검정이 된 F. asiaticum strain 18DG11과 F. graminearum strain PH-1를 비교군으로 접종하여 병원성 검정을 진행하였다. 그 결과, 각 균주별로 병원성이 다양하게 나타났으며, 72개 균주 중 71개의 균주에서 병이 발생하였다. 12개 균주는 병의 심각도가 0 이상 2 미만 사이인 약한 병원력을 나타냈고, 병의 심각도가 4 이상으로 병원력이 높은 균주는 20균주이며, 나머지 40균주는 병 심각도가 2 이상 4 미만으로 중간 정도의 병원력을 나타내는 것으로 분류되었다(Supplementary Fig. 1). 보리와 밀에서 강한 병원성을 나타낸 F. graminearum PH-1 균주의 경우 트리티케일에서는 병원성이 중간 정도로 나타냈으며, 마찬가지로 밀과 보리에서 강한 병원성이 보인 F. asiaticum 18DG11 균주는 트리티케일에서는 병원성이 나타나지 않아 균주별 병원성의 차이를 나타냈다. F. asiaticum 18DG11 균주는 같은 F. asiaticum 균주임에도 불구하고 병원성 반응이 전혀 다르게 나타나 기주 특이성 및 lineage 분석 등 추가 연구를 통해 병원성 차이에 대한 원인 구명이 필요할 것으로 판단된다. 72개의 Fusarium spp.에서 다양한 병원성 반응을 나타낸 7개 균주를 선발하여 밀에 접종하여 병원성을 비교 검정한 결과 JOS3-2 균주를 제외한 5개의 균주의 병원성이 트리티케일보다 밀에서 낮게 나타났으며, SAY1-2 균주는 밀과 유사한 병원성을 나타냈다(Fig. 3).
독소 type 분석. 트리티케일에서 분리한 91개의 곰팡이 균주 중 Penicillium spp. 등 부생균을 제외한 Fusarium spp.로 동정된 72개 균주의 독소 type을 PCR을 통해 분석하였다. 기존 붉은곰팡이 균주가 생성하는 것으로 알려진 독소 중 NIV, DON type의 독소를 분석한 결과, 72개 균주 중 68.06% 균주가 NIV를 생성하였으며, 31.94%의 균주가 DON type 독소를 생성하였다. DON의 subtype을 분석한 결과 4.17%, 27.78%의 균주가 3-acetyldeoxynivalenol, 15-acetyldeoxynivalenol 독소를 생성하는 것으로 분석되었다. 균주의 분자 동정 결과와 비교 분석한 결과, NIV type 독소를 생성하는 F. asiaticum이 트리티케일에서 분리한 균주에서 우점하는 것으로 분석되었다(Table 3). 이러한 분석 결과는 다른 작물에서 분석한 내용과 유사하다. 2010년 Ryu 등에 의한 보고에 의하면, 국내 생산 보리, 밀 등에서 발생한 붉은곰팡이 유래 독소의 분포형태를 보면 NIV 독소가 우점하고 있는 것으로 분석되었으며, 국내 생산 쌀의 독소를 분석한 결과 NIV 독소가 다른 독소에 비해 매우 높게 생성되는 것으로 분석되었다(Lee 등, 2011; Ryu 등, 2011). F. asiaticum이 생산하는 NIV type독소는 DON type 독소와 비교해 독성이 10배 이상으로 알려져 있으며, 인축에 더 유해한 것으로 보고되고 있다(Ferriogo 등, 2016). 국내에서 생산하는 쌀, 보리, 밀 등 주요 식량작물에서 발생하는 붉은곰팡이의 독소는 NIV 독소로써 그 유해성이 높기 때문에 방제가 매우 중요하며, 트리티케일의 활용성을 높이기 위해서도 트리티케일에 발생하는 붉은곰팡이병의 지속적인 모니터링 수행과 방제의 필요성을 보여주는 중요한 결과이다.
약제 내성 발생 확인. 현재 트리티케일에 등록된 농약은 20종이 있으며, 그 중 11개 약제는 제초제, 나머지 9개 약제는 절간 단축을 위한 ethephon을 주성분으로 하는 생장조정제가 등록되어 있다. 붉은곰팡이병 방제에 사용 가능한 등록 약제는 현재 없는 상태이다. 트리티케일의 붉은곰팡이병을 효율적으로 방제할 수 있는 약제를 선발하기 위해 보리와 밀에 사용 가능한 약제인 captan, fludioxonil, difenoconazole·propiconazole, hexaconazole에 대한 약제 내성 발생 여부를 확인하였다. 각 농약의 권장 농도가 함유된 배지에 균주를 치상하여 균사 생장 정도를 측정하였다. 그 결과, captan, difenoconazole·propiconazole 혼합제 및 hexaconazole에 대한 내성은 발생하지 않았다. Fusarium spp.로 동정된 72개의 균주를 대상으로 captan의 사용 권장 농도인 2,000 ppm을 넣은 배지에서 균주를 배양한 결과 52개의 균주는 균사가 전혀 자라지 않았고, 20개 균주에서는 평균 23.49±3.55 cm의 균사 생장을 보여 대조군의 반수영향생육기준치인 42.5 cm보다 작아 균사생장 억제 효과가 있는 것으로 파악되었다. Hexaconazole의 경우 20개 균주가 권장농도인 500 ppm에서 균사가 생장하였으며 그 생장 정도는 평균 19.36±4.63 cm의 생육을 나타내어 권장 농도 내에서 균사 생장억제 능력이 확인되었다.
Difenoconazole과 propiconazole 혼합제의 경우 24개의 균주가 균사 생장을 보였다. 균사 생장 정도는 평균 10.96±5.71 cm로 4가지 약제 중 균사 생장억제력이 가장 높게 나타났다. Fludioxonil의 경우 시험 균주 모두 균사 생장을 보였다. 대부분의 균주들이 균사생장이 억제되어 평균 12.46±13.71 cm의 균사 생장을 보였으나, 이 중 6개 균주의 경우 평균 균사 생장이 47.96±3.15 cm으로 약제 미처리군과 비교하여 반수영향생육 기준치인 42.5 cm의 이상으로 자라 fludioxonil에 대한 약제 저항성이 발생한 것으로 보이며, 다른 약제에 비해 균사 생장 억제 효율이 떨어진 것으로 판단된다(Table 4). Park 등(2012)에 의한 보고에 의하면 fludioxonil 처리한 붉은곰팡이 균주가 배지에 배양할 때와 포장 처리 시에도 방제 효과가 높은 것으로 보고되었다. 그 결과는 본 연구 결과와 차이를 보여 지난 10년 사이 일부 균주에서 fludioxonil에 대한 내성이 발생한 것으로 판단된다. F. asiaticum의 약제 내성 발생에 대한 보고는 fludioxonil뿐만 아니라 phenamacril, dfenoconazole, propiconazole 등의 약제에서도 보고가 되었다. 약제 내성 발생 원인은 MYOSIN5이나 CYP51 등의 유전자의 변이에 의한 것이라고 알려졌다(Li 등, 2016; Yin 등, 2009; Zhao 등, 2021). 이러한 연구결과를 토대로 트리티케일에서도 방제 효과를 높이기 위해 약제에 대한 내성 발생 모니터링과 원인 규명을 지속적으로 진행할 필요성이 있을 것으로 판단된다.
요 약
본 연구에서는 겨울철 사료작물로써 중요성이 증가하고 있는 트리티케일에서 발생한 붉은곰팡이병 원인균의 동정 및 독소 type을 분석하고 트리티케일에 효율적으로 적용 가능한 방제 약제를 선발하고자 분리된 붉은곰팡이 균주의 농약저항성 발생 여부를 분석하였다. 조성, 신조성, 신영, 신성, 세영 등 5개 트리티케일 품종에서 평균 9.15%의 병 발생 이삭률을 보였으며, 병 발생 낱알률은 29.2%로 나타났다. 그 결과 트리티케일에서 발생한 붉은곰팡이병 발생률은 평균 3.52%로 나타났다. 붉은곰팡이병에 감염된 개체로부터 총 91개의 균주가 분리되었으며, 이 중 붉은곰팡이 균주는 총 72개를 분리하였다. 분리한 균주의 형태 및 internal transcribed spacer 1, translation elongation factor 1α 유전자의 염기서열 분석 결과, 트리티케일에서 분리한 균주들은 전부 Fusarium asiaticum으로 동정되었다. 분리 동정된 균주들의 독소 type을 PCR을 통해 분석한 결과, nivalenol (NIV) type 독소는 64.6%, 3-acetyldeoxynivalenol은 4.6%, 15-acetyldeoxynivalenol은 30.8%로 대부분이 NIV type독소를 생성하는 F. asiaticum으로 분석되었다. 최근 봄철 이상 기상현상으로 여러 식량 작물에 붉은곰팡이병 발생률이 높아지고 있다. 트리티케일의 경우 붉은곰팡이병을 방제할 수 있는 등록약제가 없다. 본 실험에서는 보리와 밀에 등록된 약제를 대상으로 트리티케일에서 분리된 붉은곰팡이병 균주들의 약제 내성을 측정한 결과 captan, hexaconazole, difenoconazole·propiconazole 합제는 균사 생장 억제 효과가 있었다. Fludioxonil의 경우 72개 균주 중 6개의 균주가 권장 농도에서 반수영향생육기준치인 42.5 cm를 넘은 47.96±3.15의 균사 생장율을 보여 fludioxonil에 대한 내성이 발생한 것으로 분석되었다. 본 연구는 국내에서 처음 보고 되는 F. asiaticum에 의한 트리티케일의 붉은곰팡이병 발생 사례이며, 또한 붉은곰팡이가 생성하는 독소 타입을 구명하고, 등록 방제 약제가 없는 트리티케일에 효율적으로 방제가 가능한 내성 미발생 약제를 제시하는 중요한 연구 결과이다.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Supplement
Supplement Print
Print






