Identification of Botrytis cinerea, the Cause of Post-Harvest Gray Mold on Broccoli in Korea
Article information
Abstract
In this study, we identified the causative agent of post-harvest gray mold on broccoli that was stored on a farmers’ cooperative in Pyeongchang, Gangwon Province, South Korea, in September 2016. The incidence of gray mold on broccoli was 10-30% after 3-5 weeks of storage at 3°C. Symptoms included brownish curd and gray-to-dark mycelia with abundant conidia on the infected broccoli curds. The fungus was isolated from infected fruit and cultured on potato dextrose agar. To identify the fungus, we examined the morphological characteristics and sequenced the rDNA of the fungus and confirmed its pathogenicity according to Koch’s postulates. The results of the morphological examination, pathogenicity test, and sequencing of the 5.8S rDNA of the internal transcribed spacer regions (ITS1 and ITS4) and three nuclear protein-coding genes, G3PDH, HSP60, and RPB2, revealed that the causal agent of the post-harvest gray mold on broccoli was Botrytis cinerea. To our knowledge, this is the first report of post-harvest gray mold on broccoli in Korea.
Broccoli (Brassica oleracea var. italica), belonging to the Brassicaceae family and a native of Mediterranean regions, is one of the most popular vegetables and is distributed all over the world. Broccoli is a great source of phenolic compounds (flavonoids) and contains significant amounts of other important antioxidant chemicals such as ascorbic acid, carotenoids, selenium, glucosinolates, and sulforaphane (Conaway et al., 2000; Lin and Chang 2005; Naguib et al., 2012). These antioxidant compounds can help protect against multiple diseases, including several types of cancer, high blood pressure, macular degeneration, stomach ulcers, and type 2 diabetes (Bahadoran et al., 2013; Heber et al., 2014; Zhang and Tang, 2007). In 2013, China was the largest cauliflower- and broccoli-producing country in the world, producing 9,356,707 tons of this produce; in the same year, Korea produced 70 tons on 7 hectares of land (FAO, 2014). In Korea, broccoli is mainly cultivated in the spring and fall, but is consumed throughout the year. For summer consumption, storage of broccoli produced in the spring is necessary. However, preserving broccoli for a long time requires low temperatures and high humidity (Fernández-León et al., 2013). However, dehydration of broccoli is a major problem in low-temperature storage (Toivonen and Forney, 2004). To protect the broccoli from dehydration, water is allowed to flow on the floor during storage. This condition results in several fungal diseases.
The genus Botrytis includes over 30 species of plant pathogens with various life history traits (Fillinger and Elad, 2015). The most important species, Botrytis cinerea Pers. Fr. (teleomorph Botryotinia fuckeliana [de Bary] Whetzel), is a filamentous fungal pathogen that causes gray mold disease and significant losses in more than 200 crop species, including vegetables, fruits, and ornamental flowers both in the field and after harvest (Elad, 1997; Jarvis, 1980; Williamson et al., 2007). The pathogen is a necrotroph, triggering host cell death and causing significant damage to plant tissues, culminating in decay of the plant or harvested product. Infections with this pathogen are usually not identified at the time of harvest, but develop in a short time in humid conditions encountered during storage and transport, even at 0°C (Romanazzi et al., 2016). The taxonomy of Botrytis species has largely been based on morphological characteristics, particularly those of the mycelia, as well as the length and wide of conidiophores, conidia, and sclerotia, and species are named according to host association. These characteristics are helpful in delimiting some species, but many species are morphologically similar and growing conditions substantially affect mycelial structure (Beever and Weeds, 2007). Indeed, no key is available that can be used to identify all Botrytis species, and identification of species depending on traditional criteria can be difficult (Nielsen et al., 2002). Recently, DNA-based molecular approaches have been used widely to identify fungi, including Botrytis species. Phylogenetic analyses of Botrytis species using the sequences of three nuclear housekeeping genes and the glyceraldehyde-3-phosphate dehydrogenase gene (G3PDH), heat shock protein 60 gene (HSP60) and DNA-dependent RNA polymerase subunit II gene (RPB2) (Li et al., 2012; Staats et al., 2005; Zhou et al., 2014). Furthermore, primers targeting necrosis and ethylene-inducing protein (NEP) genes 1 and 2 can be used to identify several species of Botrytis (Grant-Downton et al., 2014; Lorenzini and Zapparoli, 2014; Staats et al., 2007). During this study, we characterized the causative agent of the post-harvest gray mold on broccoli based on morphological features, molecular phylogenetics, and pathogenicity.
Sample collection & pathogen isolation
In September 2016, signs of gray mold (disease incidence: 10-30%) were observed on broccoli after 3-5 weeks of storage at 3±1°C and 90% relative humidity in a farmers’ cooperative in Pyenonchang, Gangwon Province, South Korea. The signs observed were brownish curd and gray to dark mycelia with abundant conidia on the infected broccoli curds (Fig. 1A, Fig. 1B). To isolate this fungus, infected broccoli curds were cut into small pieces, surface sterilized with 1% sodium hypochlorite (NaOCl) for 1 min, and washed in sterile distilled water thrice and dried with sterilized filter paper. Then, the curds were placed in Petri plates containing potato dextrose agar (PDA). The plates were incubated at 20±2°C for 4 days and observed for fungal growth. For pure cultures, hyphal tips were cut after 4 days, transferred to fresh PDA, and incubated at 20±2°C for 7 days.
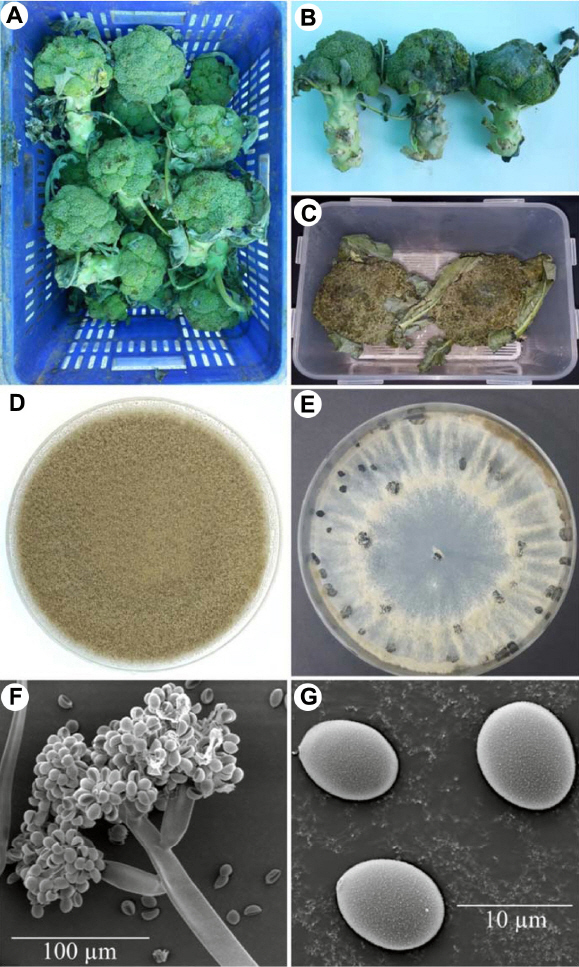
Post-harvest gray mold caused by Botrytis cinerea on broccoli. (A, B) Broccoli curds damaged by gray mold. (C) Gray mold that developed 7 days after artificial inoculation. (D) Two-week-old colony of B. cinerea on potato dextrose agar. (E) Black sclerotia on potato dextrose agar. (F, G) Conidiophore and conidia of B. cinerea under scanning electron microscope.
Growth characteristics at different temperatures
To determine the growth of this fungus at different temperatures, mycelial discs (6.5 mm in diameter) were collected from areas of active growth near the edges of 5-days-old cultures, transferred to PDA, and incubated at 1, 3, 10, 15, 20, 25, 30, and 35°C in the dark for 5 days. Three replicates were prepared. The diameters of colonies on all plates were measured using the criss-cross method (Tao et al., 2011). After 5 days of culture, the colony diameters at different temperatures were recorded.
Pathogenicity assay
For the pathogenicity test, an inoculum was prepared by harvesting conidia from 2-week-old cultures on PDA. A conidial suspension (2×106 conidia/ ml) was sprayed onto two broccolis, and another two broccolis were sprayed with sterilized water as a control. All inoculated broccoli were transferred to a plastic box that had been sprayed with sterile water on the inside to maintain high moisture content and were incubated in darkness at 20±2°C. After 5 days, dark brown lesions resembling the original signs of disease developed on the inoculated broccoli curds, whereas the control broccoli remained pristine (Fig. 1C). The pathogenicity test was carried out twice with similar results. The fungus was re-isolated from the lesions of the inoculated broccoli, and the re-isolated pathogen showed similar morphological characteristics as those of the original isolates. The fungal pathogen, therefore, fulfilled the criteria stipulated by Koch’s postulates.
DNA extraction, polymerase chain reaction (PCR) amplification, and sequence analysis
Genomic DNA was extracted using a DNeasy Plant Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s protocol. The sequences of the internal transcribed spacer and 5.8S ribosomal DNA (ITS-5.8S rDNA) and three nuclear protein-coding genes, G3PDH, HSP60, and RPB2, were amplified from total DNA by polymerase chain reaction (PCR). Primers designed for PCR amplification and sequencing are outlined in Table 1. PCR amplification was performed in a 25-µl reaction mixture containing 0.5 µl of each primer, 0.5 µl Taq DNA polymerase (Bioneer, Korea), 0.5 µl of each dNTP, 2.5 µl 10×PCR reaction buffer, 18.5 µl distilled water, and 2.0 µl template DNA. The reaction was performed in a MasterCycler Gradient thermal cycler (Eppendorf, Germany). The PCR amplification program for the ITS region consisted of an initial denaturation at 94°C for 4 min, followed by denaturation at 94°C for 1 min, annealing at 60°C for 1 min, extension at 72°C for 1 min (35 cycles), and then a final extension at 72°C for 10 min. The PCR amplification program for the RPB2 and HSP60 gene fragments consisted of an initial denaturation at 94°C for 5 min, followed by 35 cycles of denaturation at 94°C for 30 s, annealing at 55°C for 30 s, and extension at 72°C for 90 s, and then a final extension at 72°C for 10 min. For the G3PDH gene, the PCR conditions were an initial denaturation at 94°C for 5 min, 35 cycles of 94°C for 30 s, 64°C for 30 s, and 72°C for 90 s, and a final extension at 72°C for 10 min. After the PCR, DNA concentrations were estimated visually on a 1% agarose gel by comparing band intensities with that of a 100-bp DNA ladder. DNA sequencing was performed by SolGent Com. Ltd., Daejeon, Korea. The obtained nucleotide sequences were subjected to a BLASTn search in the GenBank database (http://www.ncbi.nlm.nih.gov/BLAST/). A phylogenetic analysis was carried out using MEGA6 software (Tamura et al., 2013) and the neighbor-joining method (Saitou and Nei, 1987).

Primers used for polymerase chain reaction amplification and sequencing
Identification and characterization of Botrytis cinerea
A total of five morphologically similar fungal isolates were obtained from gray molds on five broccoli samples, and of these, isolate BGM005 was used for identification. Fungal colonies on PDA at 20°C were initially white and then turned gray to dark gray with abundant conidia after 7 days (Fig. 1D). After 3 weeks, the fungus formed numerous hard, irregular, and black sclerotia ranging from 1.1-4.5×1.0-3.7 mm (n=20) near the edge of the Petri dish (Fig. 1E). Conidiophores arose singly or in groups and were straight or flexuous and septate, with an inflated basal cell colored brown to light brown and measuring 10.1–27.3×109.3–402.9 µm (Fig. 1F). The conidia were one-celled, ellipsoid, or ovoid in shape, dark brown, and measured 5.3–10.1×4.3–9.3 µm (n=50) (Fig. 1G). The growth of this fungus at different temperatures, the fungus grew fastest at 20°C and 25°C, slower at 3°C, 10°C, and 30°C, and did not grow at all at 1°C and 35°C (Fig. 2). The morphological characteristics of the identified species are summarized in Table 2. The representative isolate (BGM005) was deposited at Gangneung-Wonju National University and used for further studies.
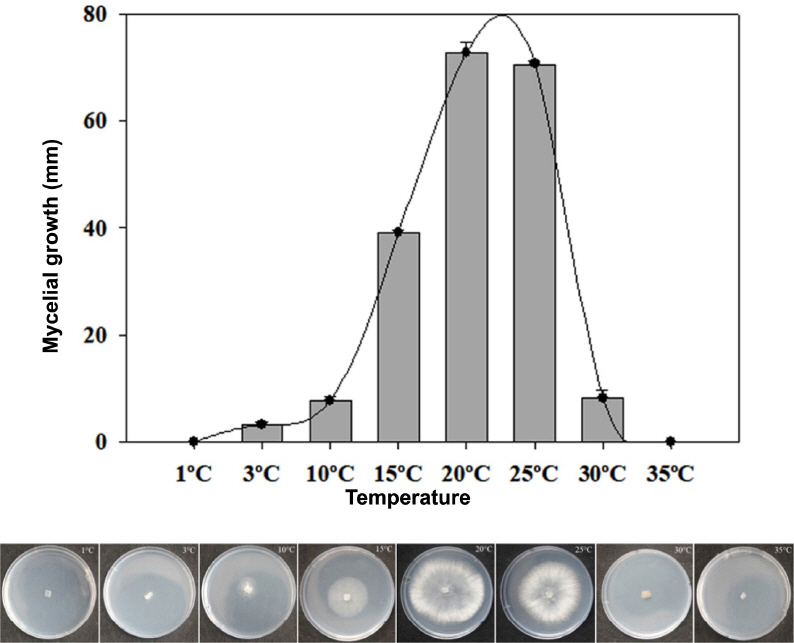
Diameters of Botrytis cinerea colonies on potato dextrose agar (PDA) after 5 days at different temperatures.

Morphological characteristics of the gray mold fungus isolated from broccoli
BLAST results for the approximately 545-bp ITS-5.8S rDNA, 756-bp G3PDH, 1028-bp HSP60, and 1093-bp RPB2 sequences from the original isolates, as well as those of the ITS-5.8S rDNA, G3PDH, HSP60, and RPB2 sequences from the reisolated fungus, were obtained, and they were 100% identical. The sequences from the representative isolate, BGM005, were deposited in the NCBI database (GenBank accession no. KY817366 for ITS-5.8S rDNA, KY817367 for G3PDH, KY817368 for HSP60, and KY817369 for RPB2). The BLAST analysis confirmed the identity of the fungus, showing 100% identity with the ITS-5.8S rDNA sequence of B. cinerea strain QT5-19 (GenBank accession no. KX822693), 100% identity with the G3PDH sequence of B. cinerea strain RSL-1 (KJ018760), 99% identity with the HSP60 sequence of B. cinerea isolate Bci-18 (KP120877), and 99% identity with the RPB2 sequence of B. cinerea strain SMGM003 (KX443704). In the phylogenetic tree based on the combined ITS-5.8S rDNA, G3PDH, HSP60, and RPB2 sequences, the representative isolate was placed within a clade comprising the reference isolates of B. cinerea and other Botrytis species (Table 3, Fig. 3).
Species of Botrytis and Sclerotinia, isolates/strains, origins, dates, and GenBank accession numbers for nucleotide sequences used in the phylogenetic analysis
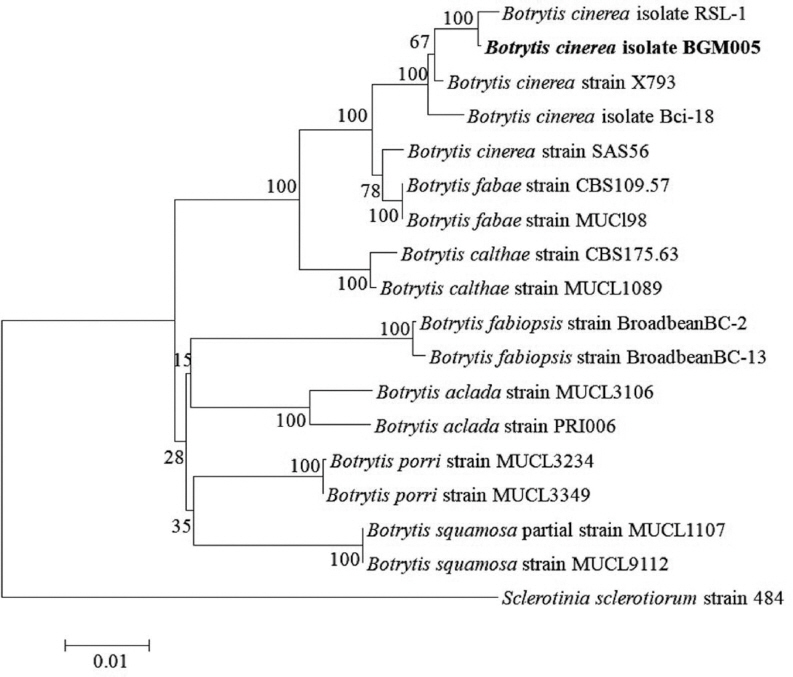
Phylogenetic analysis of Botrytis cinerea isolate BGM005, constructed using the neighbor-joining method and based on combined ITS-5.8S rDNA, G3PDH, HSP60, and RPB2 gene sequence data. Sclerotinia sclerotiorum strain 484 was used as the outgroup. The numbers at the nodes indicate bootstrap values based on 1,000 replicates. The scale bar indicates the number of nucleotide substitutions. The isolate obtained in this study is shown in bold.
Correct identification of the pathogen causing postharvest disease is central to selecting an appropriate disease control strategy. Important genera of anamorphic postharvest pathogens include Penicillium, Aspergillus, Botrytis, Fusarium, Alternaria, Colletotrichum, Dothiorella, and Phomopsis. According to a recent review, B. cinerea ranked second in a list of the top 10 fungal plant pathogens based on scientific and economic importance worldwide (Dean et al., 2012). Post-harvest decay caused by B. cinerea is of great economic importance and, in some cases, can lead to complete loss of the product. Major post-harvest losses due to B. cinerea infection occur in crops of many fruits, including apple, blackberry, blueberry, grape, raspberries, strawberry, and cherry, and vegetables, including carrot, tomato, and leafy vegetables (Droby and Lichter, 2004; Romanazzi and Feliziani, 2014). In Korea, B. cinerea has been reported as a post-harvest pathogen of blueberries, sweet cherries, sweet persimmon, carrots, and other produce (Aktaruzzaman et al., 2014, 2017a, 2017b; Kwon et al., 2011). Although this fungus has been found on broccoli in fields in Canada (Farr and Rossman, 2017), it has not been reported as a pathogen of broccoli in Korea either in the field or after harvest. Based on disease signs, morphological characteristics, and results of the phylogenetic analysis and pathogenicity test, this fungus was identified as B. cinerea Pers. (Barnett and Hunter, 1972; Ellis, 1971; Zhang, 2006). To our knowledge, this is the first report of post-harvest gray mold on broccoli in Korea. The recent finding of this disease in post-harvest broccoli suggests that gray mold may pose a serious threat to stored produce and that an effective control strategy should be developed for Korea.
Conflicts of Interest
No potential conflict of interest relevant to this article was reported.