

서론
식물 근권에 존재하는 수많은 미생물 중 식물에 유용한 Bacillus속은 식물과 직 · 간접적으로 상호작용하여 식물의 생육을 촉진하거나(Ahmad 등, 2008; Vessey, 2003), 2 차대사물질 등을 분비하여 식물병원성 미생물의 생육을 억제하는 생물학적인 방제균으로 농업적 및 환경적으로 중요한 역할을 한다(Govindasamy 등, 2010). Bacillus속은 세포벽을 분해하는 효소인 chitinase, glucanase, protease 등을 생성하고, 다양한 lipopeptide 항균물질과 volatile organic compound 등을 분비하여 식물병원균의 생육을 억제한다(Mota 등, 2017).
Bacillus속 균주에 의해 생산되는 surfactin, iturin, fengycin 같은 lipopeptide biosurfactant들은 다양한 생물학적 활성을 나타내며 동시 생성 시 상승작용으로 식물병원균에 대한 강력한 향균활성을 보여(Arima 등, 1968; Leifert 등, 1995; Montesinos, 2007; Yu 등, 2002), 식물병 방제 수단으로 정제되어 사용되고 있다(Kumar 등, 2016; Mnif 등, 2016). 특히 Bacillus amyloliquefaciens쟂ZB42는 surfactin, bacillomycin, fengycin 등의 lipopeptide들과 difficidin와 macrolactin 등의 polyketide 등 같은 항균물질들을 합성하여 식물 근권 병원성 미생물과 선충 등을 방제하는 생물적 방제 대표 균주로 알려져 있다(Chowdhury 등, 2015). 또한 Bacillus속 균주들이 생산하는 biosurfactant들에 의해서 다양한 작물병원균, Magnaporthe oryzae, Botrytis cinerea, Colletotrichum gloeosporioides, Fusarium oxysporum과 Rhizoctonia solani에 직접적인 향균활성을 보일 뿐만 아니라, 병원균 침입에 대해 작물에 유도전신저항성(induced systemic resistance)을 일으켜서 작물병 발생을 억제하는 중요한 역할을 한다고 알려져 있다(Chowdhury 등, 2015; Kim 등, 2010; Mochizuki 등, 2012; Ongena와 Jacques, 2008; Ongena 등, 2007; Park 등, 2016; Rahman 등, 2015; Yamamoto 등, 2015).
선행 연구에서 곤충의 유충에서 분리한 B. amyloliquefaciens KB3와 LM11균주는 iturin A와 surfactin 등의 lipopeptide들을 생성하여 다양한 식물병원성 곰팡이의 생육을 억제하였다(Nam 등, 2016). 특히, 두 균주가 생성하는 surfactin은 생육 단계와 배지 성분에 따라 다르고, 생물적 방제 능력이 대표적인 생물적 방제균인 Bacillus subtilis QST-713과 비슷한 수준임을 보고하였다. 일반적으로 생물적방제균 Bacillus속 균주들이 생산하는 lipopeptide 항균 물질은 강한 biosurfactant 활성을 보이고 다양한 종류의 식물병원균의 생육을 억제한다(Cao 등, 2009; Huszcza와 Burczyk, 2003). 또한 일부 그람음성균인 Pseudomonas 균주들이 생산하는 biosurfactant인 rhamnolipid가 토마토 잿빛곰팡이병균인 B. cinerea와 역병균인 Phytophthora capsici의 유주자 발아 및 균사 생장을 효과적으로 억제한다고 알려져 있다(Ahn 등, 2009; Stanghellini와 Miller, 1997). 또한 일부 생물적 방제균 Bacillus속과 Enterobacter속 균주에서 생성되는 biosurfactant들도 식물병원균 Magnaporthe grisea, Aspergillus niger, Penicillium chrysogenum, P. roqueforti와 Sclerotinia sclerotiorum의 포자를 억제하였다(Chitarra 등, 2003; Souto 등, 2004; Tendulkar 등, 2007).
생물적 방제균이 생산하는 non-ribosomal peptide synthase에 의해서 생성되는 항균 lipopeptide들은 biosurfactant의 특성상 배양여액 surface tension의 감소와 밀접한 상관관계가 있고, 생물적 방제균 선발에도 이용된다(Bodour와 Miller-Maier, 1998; Nam 등, 2016). 다양한 생물적 방제균이 생산하는 biosurfactants에 대한 연구는 주로 배양조건이나 배지에 의해 생산되는 양이나 농도에 의한 식물병원균의 항균 능력에 대한 연구가 주로 이루어져 왔다. 하지만 생물적 방제 균주의 생육단계별로 생성되는 biosurfactants의 surface tension activity와 향균활성과의 특성에 대한 명확한 관계를 나타내는 보고는 거의 없다. 따라서 본 연구에서는 다양한 식물병원균에 높은 향균활성을 보인 B. amyloliquefaciens LM11균주를 이용하여 균주의 lipopeptides가 생성되는 생육기를 결정하고, 이들 항균 물질을 함유하고 있는 세포외 상등액의 surface tension과 다양한 식물병원균과의 향균활성의 spectrum과의 상관관계를 분석하고자 하였다.
재료 및 방법
생물적 방제 균주 및 식물병원성 곰팡이 배양
본 연구에서 사용된 생물적 방제균은 장수풍뎅이 유충의 장내세포에서 분리 · 동정한 B. amyloliquefaciens LM11을 사용하였다(Nam 등, 2016). LM11균주는 tryptic soy broth (TSB; Becton Dickinson GmbH, Heidelberg, Germany) 배지에서 28°C, 120 rpm으로 5일 동안 진탕 배양하여 사용하였다. 본 연구에 사용된 식물병원성 곰팡이병균들은 토마토 시들음병균 F. oxysporum f. sp. lycopersici KACC 40032, 고추 탄저병균 C. gloeosporioides KACC 40003, 벼 잎집무늬마름병원균 R. solani AG-1 KACC 40101로 이들 균주들은 농촌진흥청 국립농업과학원 농업유전자원센터(National Agrobiodiversity Center) 미생물은행(Korean Agricultural Culture Collection, KACC)에서 분양 받아 사용하였다. 식물병원균들은 potato dextrose agar (PDA; Becton Dickinson GmbH) 배지에서 배양하였다.
향균활성 유전자 발현 및 생성 분석
LM11균주의 향균활성 능력에 관련된 biosurfactants의 유도 유전자들의 발현 양상을 quantitative real time RT-PCR (qRT-PCR)과 thin-layer chromatography (TLC) 분석을 하였다. Total RNA는 TSB 배지에서 OD600 nm=0.6 (mid-log phase), 1.8 (late-log phase), 2.2 이상(stationary phase) 배양시킨 후 NucleoZOL reagent (Macherey-Nagel, D·en, Germany)를 이용하여 회사의 protocol에 따라 분리하였다. 최종 RNA샘플은 DNase I (Qiagen Inc., Hilden, Germany)을 이용하여 정제하였다. qRT-PCR은 QuantiTect SYBR Green RT-PCR (Qiagen)을 이용하였다. 반응 샘플은 moloney murine leukemia virus reverse transcriptase (MMLV-RT; Enzynomics, Daejeon, Korea)를 이용하여 42°C에서 30분간 반응시켜 cDNA를 합성한 후 RT-PCR을 수행하였다. 사용된 ituD primers (forward, 5’-TTGAAYGTCAGYGCSCCTTT-3’; reverse, 5’-TTGAAYGTCAGYGCSCCTTT-3’, 482 bp), ituC (forward, 5’-CCCCCTCGGTCAAGTGAATA-3’; reverse, 5’-TTGGTTAAGCCCTGATGCTC-3’, 594 bp), srfA (forward, 5’-TCGGGACAGGAAGACATCAT-3’; reverse 5’-CCACTCAAACGGATAATCCTGA-3’, 201 bp)를 사용하였다(Chung 등, 2008). 16S rDNA primer는 분리 동정된 B. amyloliquefaciens LM11의 intergenic species region (ITS) 염기서열에서 program primer3를 이용하여 249 bp의 forward, 5’-GAGGAACACCAGTGGCGAAG-3’와 reverse, 5’-TAAACCACATGCTCCACCGC-3’를 제작하였다(Rozen과 Skaletsky, 2000). 증폭 반응은 cDNA가 포함된 QuantiTect RT Mix와 각 primer (10 pM)를 혼합한 총 20 µl를 최초 95°C 10분간 반응시키고, 95°C에서 30초, 60°C에서 55초, 72°C에서 30초간의 cycle을 40회 증폭시켜 각 3회 반복으로 실시하였다. Stratagene Mx3000P qPCR System (Agilent Technoloogies Inc., Santa Clara, CA, USA)과 소프트웨어를 이용하여 분석하였다. 각 유전자의 상대적인 발현량 분석을 위하여 2ΔΔ뺺T 방법을 사용하였다(Livak과 Schmittgen, 2001).
LM11균주가 생성하는 lipopeptides는 TLC 방법을 이용하였다(Arrebola 등, 2010). TSB 배지에서 28°C 배양하면서 생육기별로 n-butanol을 이용하여 추출하였다. 유기용매에 의한 분획층을 진공농축장치로 완전 농축하여 methanol로 녹인 후 TLC plate (silica gel 60; Merck, Darmstadt, Germany)에 전개시킨 후(mobile phase; chloroform:methanol:H2O=65:25:4, v/v/v) 물로 발색시켜 확인하였다(Razafindralambo 등, 1993). Reference 물질로 사용한 surfactin (CAS 24730-31-2)은 Sigma-Aldrich (St. Louis, MO, USA)로부터 구매하여 사용하였다. 또한 iturin A와 fengycin은 기존의 보고된 Rf값에 의해 검색하였으며(Arrebola 등, 2010), 총 독립적으로 3회 수행하였고 수행 시 3반복을 수행하였다.
세포배양여액의 surface tension과 향균활성 분석
B. amyloliquefaciens LM11균주는 TSB 배지에서 28°C, 120 rpm으로 15일간 배양하였다(OD600 nm=0.6 [mid-log phase], 1.8 [late-log phase], 2.2 이상[stationary phase]). LM11의 생장은 분광 광도계(UV-1601; Shimadzu, Kyoto, Japan)를 이용하여 흡광도 600 nm에서 측정하였다. 내생 포자와 영양 세포수는 배양액을 100°C에서 15분간 열 처리구와 무처리구에서 각각 생균수를 측정한 후 포자 생성을 계수하였다. 배양액의 생균수(colony forming unit)는 배양액을 10배 serial dilution 방법으로 멸균수로 희석한 후 TSB agar (Becton Dickinson GmbH) 배지에 도말하여 측정하였다. 생균수 측정은 총 독립적으로 3회 수행하였고, 수행 시 3반복을 수행하였다.
각 생육기별 배양액을 0.22 탆 필터(Millipore Filter Corp., Bedford, MA, USA) 를 이용하여 세포를 제거한 후 상등액을 사용하였다. 배양 상등액에 균주의 오염을 확인하기 위해 TSB agar 배지에 도말하여 균주의 존재 여부를 확인한 후 사용하였다. 배양여액의 경시적인 표면장력 변화는 Surface Tensiometer (K6; KRÜS GmbH, Hamburg, Germany)를 이용한 ring method를 이용하여 3회 반복 측정하였고, 총 3회를 수행하였다(Chopineau 등, 1988). 표면장력의 대조구로는 멸균수와 배지를 사용하였다.
LM11균주 배양여액의 surface tension과 향균활성 조사는 균주를 TSB 배지에서 28°C, 120 rpm으로 5일간 배양한 후 배양여액만을 상기의 방법으로 분리하여 조사하였다(Yang, 2015). B. amyloliquefaciens LM11균주 배양상등액의 surface tension 측정은 배양여액이 0%-40%가 되게 함유된 멸균된 TSB 배지 40%와 potato dextrose broth (PDB)가 60%가 되게 희석하여 각 표면장력의 값을 측정하였다. 향균활성을 측정하기 위해서는 배양여액이 0%-40%가 되게 함유된 멸균된 TSB 배지 40%와 멸균된 PDA와 혼합하여 PDB가 60%와 agar는 1.5%가 plate들을 조제하였다. Full length PDA에 5일간 배양한 공시 병원균 F. oxysporum, C. gloeosporioides, R. solani의 균사 선단에서 직경 5 mm의 균사 조각을 떼어내어 상등액 함유별로 조제된 PDA 혼합 배지 위에 치상 5일 후 균사 생육 길이를 측정하였다. 균사 생육 억제율(%)은 균주 상등액이 포함되지 않은 PDA에서 균사생육길이에 대한 각 상등액 함유 비율별로 포함된 PDA에서의 균사 생육 길이로 나타냈다. 균사 생육 억제율(%)=(대조구 균총 길이-처리구 균총 길이)/대조구 균총 길이-100. 각 처리는 독립적으로 3회 수행하였고, 수행 시 3반복을 수행하였다.
배양여액의 고추 탄저병균 포자 발아 억제
C. gloeosporioides KACC 40003을 PDA 배지에 접종하여 25°C에서 7일간 배양한 후 20 ml 멸균수를 넣어 분생포자를 수집하고 4겹의 멸균된 cheese close를 이용하여 포자현탁액을 회수하였다. 포자는 hemacytometer (Paul Marienfield GmbH & Co., Lauda-Konigshofen, Germany)를 사용하여 포자 현탁액 농도를 조절하여 사용하였다. 포자 발아율 조사는 12 well plate (SPL Life Sciences, Pocheon, Korea)에 배양여액이 0%-40%가 되게 함유된 멸균된 TSB 배지 40%와 PDB가 60%가 되게 희석하여 1 ml를 조제하였다. 조제된 배양여액에 200 µl의 포자현탁액(1×104 spores/ml)을 현탁하여 25°C에서 20시간 동안 120 rpm으로 배양하면서 발아율을 조사하였다. 광학현미경하(Leica Microsystems, Wetzlar, Germany)에서 발아관의 길이가 포자 장경의 길이 이상인 것을 발아한 것으로 계산하여 각 반복별 100개의 포자에 대한 발아 유무를 관찰하였으며 3반복으로 조사하였다. 대조구는 멸균된 TSB 배지와 60% PDB 배지를 사용하였다.
결과
균주 생육기별 항균 lipopeptide 생합성 유전자 발현과 생성
배양여액의 TLC 분석 결과, Rf 0.08-0.2인 fengycin 그룹, Rf 0.3인 iturin 그룹과 Rf 0.7-0.75인 surfactin 그룹들이 LM11균주의 배양여액에서 관찰되었다. 이러한 항균 lipopeptide들의 생성량은 late log와 정지기의 배양여액에서 대량으로 검출되었지만, 지수기의 배양여액에서는 소량이 검출되었다(Fig. 1A).
Fig. 1
Transcriptional and biochemical analysis of Bacillus amyloliquefaciens LM11 biosurfactant productions. (A) Lipopeptide fractions from B. amyloliquefaciens LM11 were separated on thin-layer chromatography plate (silica gel 60) and the relative distance (Rf) of each lipopeptide are indicated. (B) Quantitative real time RT-PCR analysis was performed twice using primers specific to ituC, ituD, and sfrA genes, and the expression levels were normalized against 16s rRNA gene levels. Relative transcript quantification was performed using the 2-ΔΔCT method. The values represent mean±standard error of triplicates of three independent experiments. Data were analyzed through one-way ANOVA (P<0.05), and if the F test was significant, differences were further elucidated through Duncan’s multiple range test. Different letters indicate significant different at P<0.05. (C) Antifungal activities of biosurfactant fractions extracted from different growth phases of B. amyloliquefaciens LM11 against phytopathogenic fungal pathogens. The cell-free supernatants were acidified and extracted into n-butanol, and dissolved with methanol. The paper discs were loaded with a partial purified from LM11 grown different growth phases. The image is represented three independent experiments with similar results. Bacterial cells were grown in tryptic soy broth (TSB) broth at 28°C with shaking at 120 rpm. Total RNAs and lipopeptides were extracted at defined growth.
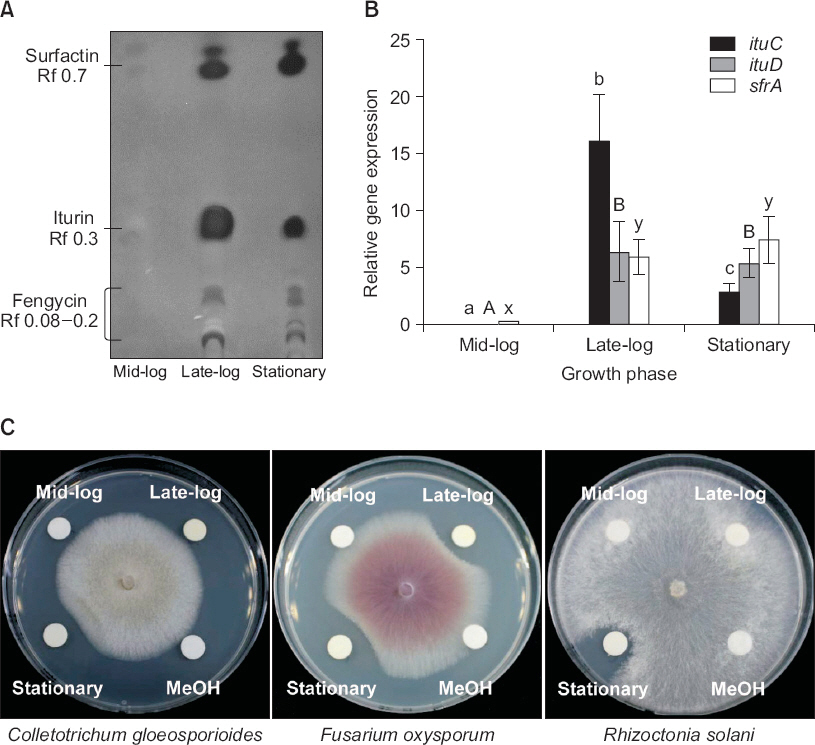
항균 lipopeptides의 생성 시기와 이들 물질의 생합성 유전자의 발현 시기가 동일하였다(Fig. 1B). LM11균주의 항균 lipopeptides들의 유전자 발현은 mid-log기의 LM11균주에 비해 late-log와 정지기에서 통계적으로 유의하게 높았다(Fig. 1B). Iturin과 surfactin의 생합성에 관련된 ituC, ituD와 sfrA 유전자들의 발현도 late-log phase와 정지기에서 유의적으로 높았다(Fig. 1B). 흥미롭게 iturin은 정지기보다 late-log기에서 통계적으로 유의하게 높았다.
LM11균주 생육기별 배양여액의 식물병원성 곰팡이에 대한 향균활성도 late-log와 정지기에서 추출한 배양여액에서는 다양한 식물병원성 곰팡이의 균사생육억제 효과를 보였지만, 지수기 배양여액은 균사생육억제 능력을 보이지 않았다(Fig. 1C).
배양여액의 surface tension과 향균활성과의 상관관계
LM11균주 배양여액의 surface tension은 mid-log기(평균 34 mN/m)보다 late-log기와 정지기(평균 30 mN/m)에서 유의하게 낮았다(Table 1). 대조구로 사용된 멸균수와 TSB 배지의 surface tension값은 73 mN/m였다. LM11균주의 정지기 배양여액(30 mN/m)은 C. gloeosporioides, F. oxysporum과 R. solani의 균사생육 억제 능력이 평균 24%였다(Fig. 1C).
Table 1
Cell density and biosurfactant activity of lipopeptides at different growth phase of Bacillus amyloliquefaciens LM11
| Growth phase | Vegetative cell (log cfu/ml)* | Endospore (log cfu/ml)† | Surface tension of culture fluids (mN/m)‡ |
|---|---|---|---|
| Mid logarithmic | 4.12±0.77 a | - | 34.2±0.52 b |
| Late logarithmic | 8.33±0.31 b | 1.12±1.33 a | 30.0±0.12 a |
| Stationary | 9.66±1.09 c | 2.95±6.60 a | 30.2±0.06 a |
The values represent mean±standard error of triplicates of three independent experiments.
Data were analyzed through one-way ANOVA (P<0.05), and if the F test was significant, the differences were further elucidated through Duncan’s multiple range test. Different letters indicate significant different at P<0.05.
* Cells of B. amyloliquefaciens LM11 were grown in tryptic soy broth (TSB) to mid log-phase (OD600 nm=0.8), late log-phase (OD600 nm=1.8), and stationary-phase (OD600 nm>2.2) at 28°C. Colony forming unts of B. amyloliquefaciens LM11 were determined by preparing 10-fold serial dilutions and 0.1 ml from each dilution was inoculated into TSB agar plates.
LM11균주의 정지기 배양여액의 다양한 식물병원균에 대한 균사생육억제 능력이 배양여액의 농도와 직접적인 연관이 있었다. 즉 배양여액 40% 함유된 PDA 배지에서 공시한 병원성 곰팡이의 균사 생육 억제는 75.3%-89.7%로 강한 향균활성을 나타냈다(Fig. 2). 배양여액 20%는 식물병원균의 균사 생육을 50% 이상 억제하였지만, 20% 이하에서는 균사생장억제 능력이 현저히 감소하였다.
Fig. 2
Effects of the cell-free supernatant of Bacillus amyloliquefaciens LM11 grown on inhibition mycelial growth or spore germination of phytopathogenic fungi. (A-C) Cell-free supernatants from B. amyloliquefaciens LM1 grown stationary phase showing antifungal activity against phytopathogenic fungi. The fungal mycelial growth inhibitions of the LM11 strain were determined on potato dextrose agar amended with different concentrations of cell-free stationary phase culture supernatants of LM11. (A) Fo, Fusarium oxysporum f. sp. lycopersici KACC 40032, (B) Cg, Colletotrichum gloeosporioides KACC 40003, (C) Rs, Rhizoctonia solani AG-1 KACC 40101. (D) Conidia germination were determined in 1/3 potato dextrose broth (PDB) medium in different concentration of cell-free supernatant of LM11. The spore germination was determined under light microscope after 24 h of incubation with at least 100 spore/experiment. Spores which produced germ tubes longer than their diameter were considered to have germinated. Error bars presented are mean±standard error of triplicates of three independent experiments. Data were analyzed through ANOVA (P<0.05), and if the F test was significant, differences were further elucidated through Duncan’s multiple range test. Different letters indicate significant different at P<0.05. Linear regressions where X is the independent variable concentration of cell-free supernatant of the LM11 stationary phase culture and Y is the dependent variable spore germination inhibition of C. gloeosporioides were obtained. We calculated the Spearman correlation coefficient (R-value) and its corresponding P-value.
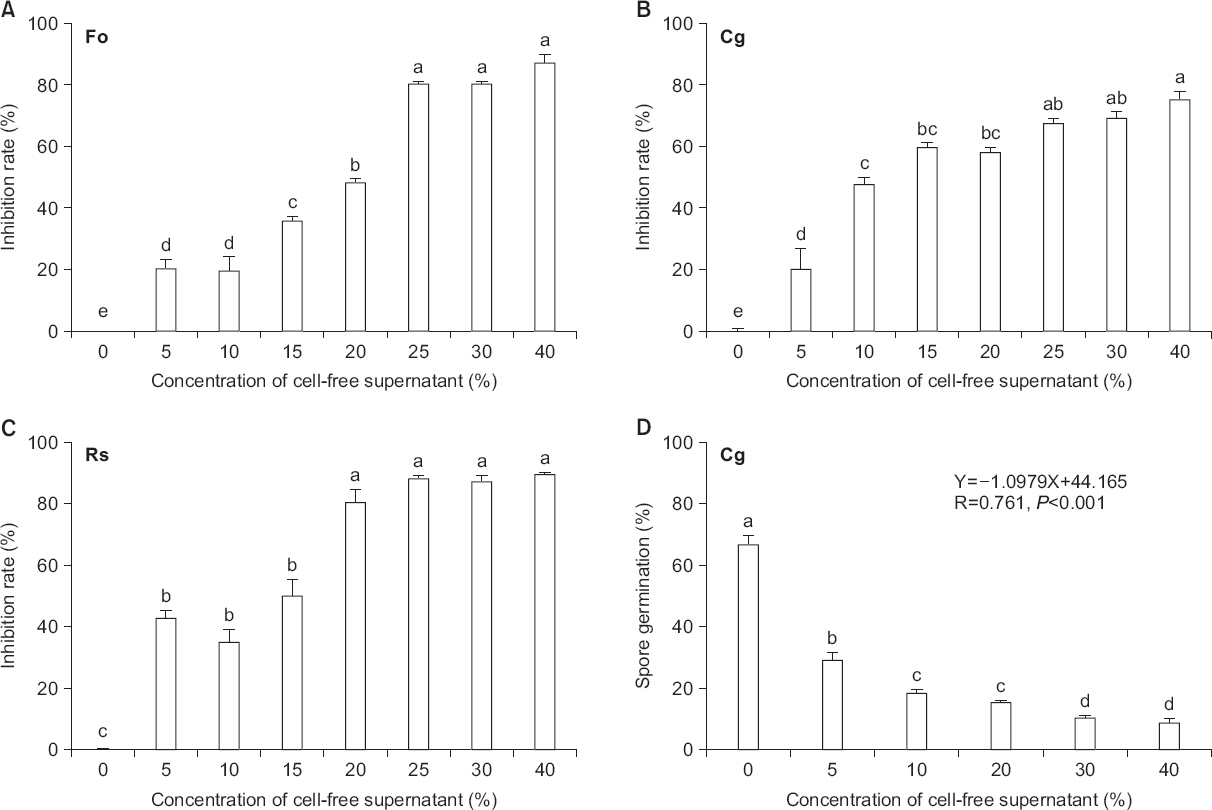
LM11균주의 정지기 배양여액 함량이 높을수록 C. gloeosporioides 포자발아율과 부의 상관관계가 있었다(R=0.761, P<0.001; Fig. 2D). 배양여액 30% 함유된 처리구에서는 10%로 가장 낮은 발아율을 나타냈다.
LM11균주 정지기 배양여액 surface tension과 식물병원성 곰팡이균주의 균사생육억제와의 상관관계 분석결과, C. gloeosporioides, F. oxysporum과 R. solani에 대해 각각 R=0.973**, 0.951**, 0.977**로 0.01% 수준에서 고도로 유의하였다(Fig. 3). C. gloeosporioides와 R. solani의 경우 surface tension이 42.0 mN/m와 51.8 mN/m인 정지기 배양여액 10%에서 약 50% 이상의 균사생육억제 능력을 나타냈지만, F. oxysporum은 37.1 mN/m (30%)에서 50% 이상의 향균활성 능력을 나타냈다.
Fig. 3
Correlation between the levels of surface tension and the antifungal activity of the Bacillus amyloliquefaciens LM11 bacterial cultures grown in stationary phase. (A) Fo, Fusarium oxysporum f. sp. lycopersici KACC 40032, (B) Cg, Colletotrichum gloeosporioides KACC 40003, (C) Rs, Rhizoctonia solani AG-1 KACC 40101. The cell-free culture of stationary phase was serially diluted (0%-40%) with sterile tryptic soy broth (TSB) broth and the surface tension was measured. Mycelium growth inhibitions of the different concentrations of the LM11 culture filtrates against phytopathogenic fungal pathogens were examined by placing 5 mm mycelium agar disc (taken from 5 days old potato dextrose agar [PDA] plates) at the center of PDA plates containing different concentrations of the cell-free culture supernatants of LM11 grown stationary phase. All plates were incubated at 28°C for 5 days. Fungal growth was observed as an increase in colony diameter. Three replicate plates were used per assay. Percentage mycelium inhibition rate of the LM11 culture filtrate was calculated by the formula: Mycelium growth inhibition (%)=(A-B)/A×100, where A is mycelial growth in the control PDA and B is mycelia growth in the amended PDA plates. Each point represents the means of different diluted cell-free supernatants of B. amyloliquefaciens LM11 grown in TSB until stationary phase shown with three to five replicates. Data from three independent replicate trials were subjected to statistical analysis. Linear regressions where X is the independent variable surface tension and Y is the dependent variable mycelium growth inhibition were obtained. We calculated the Spearman correlation coefficient (R-value) and its corresponding P-value.

고찰
생물적 방제능력을 갖는 Bacillus속 균주는 ribosome에 의해 합성되는 항생물질(Kolter와 Moreno, 1992)과 non-ribosomal peptide synthase가 관여하여 생성되는 항생물질을 동시에 생산한다고 알려져 있어 다양한 식물병원균의 생물적 방제제로 연구되고 있다(Katz와 Demain, 1977; Kolter와 Moreno, 1992; Sieber와 Marahiel, 2003; Stein, 2005). 이러한 Bacillus속 균주가 생산하는 surfactin, fengycin, iturin 같은 lipopeptide들은 non-ribosomal peptide synthase에 의해 생합성되는 대표적인 항생물질로서 biosurfactant로서 target 병원균의 세포막을 파괴하여 병원균의 생육을 억제한다고 알려져 있다(Peypoux 등, 1999; Romero 등, 2007; Touré 등, 2004). 이러한 항균 lipopeptide들의 생성량이 향균활성과 직접적인 관련이 있다고 알려져 있다. B. subtilis CMB32의 배양여액에 생산된 iturin A, fengycin과 surfactin A와 같은 항균 lipopeptide들이 식물병원균 C. gloeosporioides를 효과적으로 방제하는 주 물질로 작용한다고 알려져 있다(Kim 등, 2010; Ongena와 Jacques, 2008). 또한 B. amyloliquefaciens FZB42, B94, KB3와 S13-3의 균주가 생산하는 surfactin, fengycin, iturin A, bacillomycin D와 같은 항균 lipopeptide들이 다양한 식물병원성 곰팡이의 생육 억제에 주 물질들로 보고되었다(Chowdhury 등, 2015; Mochizuki 등, 2012; Nam 등, 2016; Yu 등, 2002). B. subtilis SPB1이 분비하는 crude lipopeptide mixture의 R. solani에 대한 MIC lipopeptide 농도는 4 mg/ml였다(Mnif 등, 2015).
본 연구에 사용된 B. amyloliquefaciens LM11균주의 TLC 분석과 유전자 발현 분석에 의해 기존에 보고된 fengycin, iturin과 surfactin 그룹(Arrebola 등, 2010; Romero 등, 2007)들이 LM11균주의 주 항균물질임을 밝혔다. 또한, LM11균주는 surfactin, iturin, fengycin 같은 항균 lipopeptide들이 생육초기보다는 late-log기부터 유전자가 발현되고 생성되어 이들 물질을 함유한 배양여액이 식물병원균의 생육을 억제한다는 것을 의미한다. 일반적으로 생물적방제 Bacillus속 균주들의 향균활성은 지수기 세포에 비해 정지기 세포에서 높다(Slepecky와 Hemphill, 2006; Yoshida 등, 2001). 생물적 방제균 Bacillus속 균은 한 가지 이상의 필수 영양 성분이 고갈되면 정지기가 되며, 내생포자 형성이 시작되고, 세포 외부로 다양한 분해 효소와 이차대사물질의 생합성에 관련된 유전자들의 발현이 유도되고 분비된다(Marahiel 등, 1993). 정지기에서의 이러한 다양한 반응들은 영양 및 환경 스트레스에 대한 생존 전략의 하나이며 항생물질 생산은 생물적 방제 균주로서 제한된 양분의 환경에서 경쟁력을 강화하기 위한 수단으로 생각된다.
생물적 방제 Bacillus속 균의 배양여액 surface tension은 항균 lipopeptide 생성량과 밀접한 관계가 있다. Bacillus속의 배양여액 surface tension이 mid-log기에서 30-40 mN/m로 감소하였고(Kim 등, 2000; Lee 등, 2002; Singh 등, 2014; Varadavenkatesan와 Murty, 2013), late-log기와 초기 정지기 배양여액에서 항균 lipopeptides의 생산이 증가되어 가장 낮은 surface tension을 보이는 결과와 유사하였다(Lee 등, 2002; Mulligan 등, 1989).
LM11도 향균활성을 나타내는 lipopeptide 함량(배양여액 surface tension)과 식물병원균에 대한 향균활성과 강한 상관관계를 보임을 증명하였다. 식물병원성 곰팡이에 대한 향균활성과 LM11의 배양여액 surface tension과의 관계는 완전 부의 상관관계를 보였다(R=0.951-0.977, P<0.001). 또한 각 병원균에 대해 50% 이상의 효과적인 다양한 식물병원곰팡이의 균사 생육억제능력을 얻기 위해서는 최소 정지기 배양여액 surface tension이 38 mN/m 이하임을 밝혔다. 이러한 결과는 항균 lipopeptide를 생성하는 생물적 방제균의 배양여액 surface tension값을 식물병원균의 억제 능력을 판단할 수 있는 지표로서 가능성을 제시하였다. 하지만, 이들 배양여액 surface tension값은 균주의 생육기에 따라 향균활성이 다르기 때문에 차후 생육기별 배양여액 surface tension에 대한 향균활성 spectrum과 정지기에서 항균 lipopeptide 이외에 다른 향균활성 인자가 작용하는지 등에 연구가 필요할 것으로 생각된다.
요약
장수풍뎅이 유충의 장내세포에서 분리한 Bacillus amyloliquefaciens LM11은 surfactin, iturin, fengycin 같은 biosurfactants lipopeptide를 생산하여 식물병원성 곰팡이의 성장을 강하게 억제하였다. LM11균주 성장단계에 따라 biosurfactant 생산과 surface tension은 상당히 유의한 차이가 있었다. 항균 물질인 surfactin, iturin, fengycin의 생합성 유전자는 정지기에 도달하면서 집중적으로 발현되었고 그 생산량도 높았다. 또한 LM11균주를 제거한 배양 상등액 함량의 농도에 따라 고추 탄저병원균의 포자발아와 높은 부의 상관관계가 있었다(R=0.761, P<0.001). 식물병원성 곰팡이의 균사생장억제를 위한 최소 surface tension 수준은 38.5 mN/m였다(R=0.951-0.977, P<0.001). 본 연구 결과는 B. amyloliquefaciens LM11의 biosurfactant가 식물병에 대한 생물학적 방제에 중요한 항진균 대사물질로 작용하며, 배양액의 surface tension 측정은 생물학적 방제제의 최적 사용을 위한 기초 지표로 사용될 수 있음을 보여 주었다
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






