



Introduction
Quercus (oak) is an important genus throughout the forests of the Zagros and has been in the area for approximately 5,000 years (Mollaei, 2019). Persian oak (Quercus brantii Lindl) as the main tree species covers nearly five million hectares of the Zagros Mountains, from the southwest to the north-west of Iran (Sabeti, 1994). Granted the role of oak trees in maintaining water supplies and in the mitigation of climate change (Mollaei, 2019), any reduction and mortality of oak trees is considered a significant threat to the ecosystems (Henriques et al., 2012; Linaldeddu et al., 2014).
Charcoal canker disease, which could affect both young and adult trees, was first reported in western forests of Iran in 2011, Zagros Mountains in Kermanshah, Ilam, and Lorestan provinces, on the species of Zelkova carpinifolia (Pall.) K. Koch, Quercus castaneifolia C. A. Mey and Q. brantii (Mirabolfathy et al., 2011). Symptoms of this disease include discoloration of infected tissues, browning of leaves, and dieback of branches of part/all of the tree in one growing season. Furthermore, a large number of black stromata with the sexual reproductive organs (perithecia) are observed as a black carbonaceous layer on the stem surface (Franceschini and Luciano, 2009). In addition to Iran, charcoal canker disease of oak has been reported in various countries in Europe, New Zealand, North America, and Asia (Jurc and Ogris, 2006; Mirabolfathy et al., 2011; Vasilyeva et al., 2007).
Biscogniauxia mediterranea (de Not.) Kuntze. (Torres 1985) and Obolarina persica (Mirabolfathy et al., 2013), were introduced as pathogens responsible for charcoal canker in the inner bark of hardwood species such as cork oak. These species spend part of their lifetime as an endophyte in infected trees that are asymptomatic (Vannini et al., 2009) and can act as an opportunistic pathogen and attack weakened hosts under environmental stresses such as drought stress and high abnormal temperature (Linaldeddu et al., 2011; Ragazzi et al., 2001).
B. mediterranea has been introduced as one of the causes of decline in oak tree species in North America, Europe, Asia, Africa, and New Zealand (Collado et al., 2001; Giambra et al. 2009; Jurc and Ogris, 2006). Moreover, other species of the genus Biscogniauxia infect oak trees, including B. uniapiculata (Penz. and Sacc.) Whalley and Laessoe, B. nummularia (Bull.) Kuntze and B. nothofagi Whalley, Laessoe, and Kile, have been reported to cause cankers in other tree species, such as Nothofagus, Fagus, and Eucalyptus (Nugent et al., 2005).
It is worth mentioning that there are some studies con-cerning occurrence and outbreak of charcoal canker disease on different species of oak in different countries of the world such as Russia, Africa, Europe, North and Central America, the Mediterranean basin (La Porta et al., 2008), Portugal (Henriques et al., 2014), and Slovenia (Jurc and Ogris, 2006). However, there is little information documented on the effects of deficit and full irrigation on the oak trees in Iran, particularly in southern Iran, forests of the South Zagros, Kohgiluyeh & Boyer-Ahmad and Fars provinces, despite the wide geographical distribution of Quercus brantii Lindl and also the existence of rich and unique Persian oak germplasm in Kohgiluyeh & Boyer-Ahmad province. According to local studies, charcoal canker disease has spread in Zagros forests over the past decade, due to climate changes including reduced rainfall about 35% (Emadodin et al., 2019; Mansouri Daneshvar et al., 2019) and drought stress (Mirabolfathy et al., 2011). The present study was, therefore, designed and implemented in two sections: the first section refers to the identification of fungal pathogens causing charcoal canker disease in the oak forests of South Zagros i.e., Kohgiluyeh & Boyer-Ahmad and Fars provinces based on morphological characteristics and molecular assay using phylogenetic analysis of the DNA sequence data from β-tubulin, α-actin genes, and ITS (internal transcribed spacer)-rDNA regions, and the second section focuses on disease symptoms’ evaluation via pathogenicity test in non-inoculated or inoculated Q. brantii seedlings under moisture environments (control and intense stress) to evaluate the main effects of environments, charcoal canker disease agent isolates, oak ecotypes and specifi-cally their dual (environment×isolate, isolate×ecotype and environment×ecotype) and triple interaction effects (environ-ment×isolates×ecotype).
Material and Methods
Sampling and fungal isolation.
From early September to late March 2018-2019, surveys were conducted to identify charcoal canker disease at eight oak forests located in the Zagros Mountains (southern Iran): five regions in Kohgiluyeh & Boyer-Ahmad province (Basht, Gachsaran, Dehdasht, Yasouj, and Sisakht), and three regions in Fars province (Rostam, Mmasani, and Kazerun) (Supplementary Fig. 1). The samples of black stromata containing perithecia were collected from symptomatic parts of each oak tree (Fig. 1), taken to the laboratory in polyethylene bags and kept in the refrigerator (4°C) until diagnostic lab isolation were performed. The number of samples collected in each region varied according to the extent of disease spread and symptoms.
Fig. 1.
The symptoms of charcoal canker on Quercus brantii: dieback and death of the oak trees (A), the production of carbonaceous stroma in the trunk (B, C), dark exudation in the trunk (D, E), and the oak bark scaling (F).

In order to isolate oak charcoal canker disease agent, the infected samples were cut into 5-10 cm segments with a sterile scalpel, sterilized in 70% ethanol for 60 sec, rinsed in sterile deionized water, placed in glass Petri dishes, and kept under high humidity. They were then examined daily until the ascospores were released from the stromata surface. The ascospores were spread by a sterile loop on 2% water-agar medium and incubated at 25°C. After 24 hr, single-germi-nated ascospores were transferred to PDA (potato dextrose agar; 12 g/l potato extract, 10 g/l dextrose, 1.2% agar, with 50 µg of modified kanamycin to prevent bacterial contamination) for recording growth rates and cultural characteristics.
Morphological identification.
To identify the fungal species, all collected fungal isolates were morphologically evaluated and classified in accordance to genus and species de-scriptions in valid taxonomic references (Ju and Rogers, 2001; Ju et al., 1998; Mirabolfathy et al., 2013). For this purpose, the experiment was arranged in a completely randomized design that was repeated three times. Thus, fungal isolates were first cultured in a variety of media including PDA, malt extract agar, and potato carrot agar. Afterward, indices such as colony morphology, cultural characteristics (growth rate, growth temperatures, and diameter of the hyphae) as well as morphology of fungal structures including teleomorph (stroma, perithecium, ascus, and ascospore) and anamorph (conidia and conidiophores) were determined. In order to induct conidia and conidiophores (if required), pure culture of fungal isolates in PDA medium under light/dark conditions for 12 hr with a mixture of fluorescent light and near-ultraviolet light at 25±2°C for 1 week was placed. The purified isolates were then stored on lyophilized filter paper strips at -40°C. Morphological features were evaluated with the Olympus DP72 camera on the Olympus BX51 microscope (Tokyo, Japan). Measurements were performed with the Cell Sense Entry measurement module. Based on at least 50 microscopic measurements, the dimensions of each fungal structure were calculated.
DNA extraction and sequencing.
A molecular trial was conducted in the present study following initial morphological assay of fungal isolates, in which 20 isolates from Kohgiluyeh & Boyer-Ahmad (Iran 1 to Iran 10) and Fars provinces (Iran 11 to Iran 20) were randomly selected (Table 1). After DNA extraction, sequencing of different gene regions (i.e., ITS-rDNA, α-actin, and β-tubulin), polymerase chain reaction (PCR) reaction, etc. phylogenetic analysis, and molecular identification of fungal isolates were performed. To this end, the following steps were designed and implemented: first, the selected isolates were cultured in potato dextrose broth medium for 5 days at 25°C on a rotary shaker; the mycelium was then collected and freeze-dried (pure cultures). Then, observing the CTAB protocol as described by Murray and Thompson (1980), genomic DNA of pure cultures was ex-tracted. The quality of the genomic DNA was assayed using 1.1% agarose gel, visualized by staining with ethidium bromide solution, and the genomic DNA was stored at −20°C until further analysis. The pair of primers ITS1/ITS4 (White et al., 1990), T1 (O'Donnell and Cigelnik, 1997)/Bt2b (Glass and Donaldson, 1995), and ACT-512F/ACT-783R (Carbone and Kohn, 1999) were used to amplify ITS-rDNA regions, α-actin and β-tubulin genes, respectively.
Table 1.
Biscogniauxia isolates used in phylogenetic analyses in this study
| Species name | Isolate | Collecting data | GenBank accession no. | ||
|---|---|---|---|---|---|
| ITS-rDNA | α-Actin | β-Tubulin | |||
| Biscogniauxia mediterranea | Iran1 (Iran-G1) | Present study | MW287382 | MZ191092 | MZ191087 |
| B. mediterranea | Iran2 (Iran-G25) | Present study | MW287383 | MZ191093 | MZ191088 |
| B. mediterranea | Iran3 (Iran-S37) | Present study | MW287384 | MZ191094 | MZ191089 |
| B. mediterranea | Iran4 (Iran-S39) | Present study | MW287385 | MZ191095 | MZ191090 |
| B. mediterranea | Iran5 (Iran-B43) | Present study | MW287386 | MZ191096 | MZ191091 |
| B. mediterranea | Iran6 (Iran-B53) | Present study | MZ769369 | MZ782155 | MZ782170 |
| B. mediterranea | Iran7 (Iran-D53) | Present study | MZ769372 | MZ782156 | MZ782171 |
| B. mediterranea | Iran8 (Iran-D56) | Present study | MZ769381 | MZ782157 | MZ782172 |
| B. mediterranea | Iran9 (Iran-Y59) | Present study | MZ769375 | MZ782158 | MZ782173 |
| B. mediterranea | Iran10 (Iran-Y60) | Present study | MZ769378 | MZ782159 | MZ782174 |
| B. mediterranea | Iran11 (Iran-R 61) | Present study | MZ769370 | MZ782160 | MZ782175 |
| B. mediterranea | Iran12 (Iran-R65) | Present study | MZ769373 | MZ782161 | MZ782176 |
| B. mediterranea | Iran13 (Iran-R70) | Present study | MZ769382 | MZ782162 | MZ782177 |
| B. mediterranea | Iran14 (Iran-M74) | Present study | MZ769376 | MZ782163 | MZ782178 |
| B. mediterranea | Iran15 (Iran-M86) | Present study | MZ769379 | MZ782164 | MZ782179 |
| B. mediterranea | Iran16 (Iran-M91) | Present study | MZ769371 | MZ782165 | MZ782180 |
| B. mediterranea | Iran17 (Iran-M105) | Present study | MZ769374 | MZ782166 | MZ782181 |
| B. mediterranea | Iran18 (Iran-K112) | Present study | MZ769383 | MZ782167 | MZ782182 |
| B. mediterranea | Iran19 (Iran-K115) | Present study | MZ769377 | MZ782168 | MZ782183 |
| B. mediterranea | Iran20 (Iran-K117) | Present study | MZ769380 | MZ782169 | MZ782184 |
| B. anceps | BCRC34029 | Rogers et al. (1996) | EF026132 | AY951783 | AY951671 |
| B. arima | BCRC34030 | Ju et al. (1998) | EF026150 | AY951784 | AY951672 |
| B. atropunctata | YMJ 128 | Ju et al. (1998) | JX507799 | AY951785 | AY951673 |
| B. capnodes | BCRC34032 | Ju et al. (1998) | EF026131 | AY951787 | AY951675 |
| B. citriformis | BCRC34034 | Ju et al. (1998) | JX507801 | AY951789 | AY951678 |
| B. cylindrispora | BCRC33717 | Ju and Rogers (2001) | EF026133 | AY951791 | AY951679 |
| B. formosana | BCRC33718 | Ju and Rogers (2001) | JX507802 | AY951792 | AY951680 |
| B. granmo | BCRC34035 | Ju et al. (1998) | JX507803 | AY951793 | AY951681 |
| B. latirima | BCRC33729 | Ju and Rogers (2001) | JX507804 | AY951794 | AY951682 |
| B. latirima | BCRC34036 | Hsieh et al. (2005) | EF026135 | AY951795 | AY951683 |
| B. mediterranea | BCRC34037 | Ju et al. (1998) | EF026134 | AY951796 | AY951684 |
| B. nummularia | H86 | Pazoutová et al. (2012) | GQ428318 | GQ428312 | GQ428324 |
| B. simplicior | BCRC34038 | Hsieh et al. (2005) | EF026130 | AY951798 | AY951686 |
| B. uniapiculata | BCRC34039 | Hsieh et al. (2005) | JX507805 | AY951799 | AY951687 |
| B. philippinensis | BCRC33720 | Ju and Rogers (2001) | EF026136 | AY951797 | AY951685 |
| Hypoxylon rubiginosum | BCRC34116 | Ju and Rogers (2001) | EF026143 | AY951862 | AY951751 |
| Obolarina persica | BCRC34960 | Mirabolfathy et al. (2011) | JX507807 | JX507798 | JX507796 |
The PCR reaction mixtures in 20 μl volume were prepared containing 0.04 μM of each primer (Microsynth, Switzerland), 0.4 μM dNTPs (MBI Fermentas, Germany), 1× Dream Taq Buf-fer (MBI Fermentas), and 0.4 U Dream Taq DNA polymerase (MBI Fermentas). The PCR conditions were as follows: primary denaturation step for 6 min at 96°C followed by 35 cycles of 30 sec at 95 °C, 30 sec at 52°C (ITS and act) or 58°C (βtub) and 1 min at 72°C, with a final extension of 8 min at 72°C. The amplicons were analyzed on a 1% agarose gel then stained with ethidium bromide, and viewed under UV light to check the amplification size and purity (GelDoc, Bio-Rad Laborato-ries, Hercules, CA, USA). The PCR products were purified by a PCR purification kit (Fermentas, UK), and sequencing was performed in line with the method described in Hsieh et al. (2005, 2009) by Macrogen Co. (Seoul, Korea).
Phylogenetic analysis.
βtub, act, and ITS-rDNA sequences were blasted separately using Megablast to identify their closest neighbors. βtub, act, and ITS-rDNA sequences generated in the present study, combined with sequences of rep-resentative taxa from GenBank (Table 1), to determine the taxonomic status of the Biscogniauxia species. New nucleotide sequences were deposited in GenBank (http://www. ncbi.nlm.nih.gov), and the accession numbers of sequences are given in Table 1. The forward and reverse sequences of βtub, act, and ITS-rDNA were aligned initially by program Sequencer ver. 5.1 (Gene Code). Final alignments were performed using Geneious version 7 (Biomatters, Auckland, New Zealand). The most appropriate model of sequence evolution was evaluated for each dataset with JModeltest v.2.1.4 using the Bayesian information criterion (BIC) for the following phylogenetic analyses. Multi-locus phylogenetic Bayesian inference analyses were done with MrBayes v3.2.2 (Ronquist et al. 2012). act, βtub, and ITS sequences were di-vided into three partitions. Four Markov chains were run for 10,000,000 generations, with a burn-in fraction set to 0.25. Maximum parsimony (MP) phylogenies were estimated using heuristic searches in PAUP ver. 4.0a133 (Swofford, 2002) with bootstrap analysis of 1000 replicates to test the support of the branches. Hypoxylon rubiginosum (BCRC34116) is a close species to B. mediterranea and is used as an outgroup taxon to root phylogenetic trees for Biscogniauxia species (Mirabolfathy et al., 2011).
Pot experiment and pathogenicity test.
Two-year-old potted seedlings of Q. brantii were used. Briefly, from oak forests of five regions of Kohgiluyeh & Boyer-Ahmad province, oak acorns were collected separately from healthy Persian oak trees (70 random oak acorns in each region/ecotype, 560 samples). Then the acorns were sown in plastic pots containing forest soil and maintained for 24 months in the greenhouse of the Faculty of Agriculture, Yasouj University under normal irrigation conditions, 25-35±2°C and 55±5% humidity (each pot contains one seedling).
The treatment combinations in the pot experiment included two levels of irrigation regime, two levels of inoculation with two types of isolates, and five levels of ecotype/region. Regarding the isolates, of the 120 fungal isolates that were isolated from infected tissues of Q. brantii trees of Kohgiluyeh & Boyer-Ahmad and Fars provinces forests, two fungal isolates “Iran-G 1” and “Iran-M 70” were selected (Supplementary Table 1). Indices such as growth rate and type of disease-causing species were considered in the selection of isolates.
After 24 months of growing under greenhouse conditions, 480 two-year-old potted seedlings of Q. brantii (60 seedlings per ecotype) were inoculated with both fungal isolates. As a result, half of the seedlings per ecotype (n=30) were inoculated by Iran-G1 isolate and the other half seedlings by Iran-M70 isolate. Eighty non-inoculated seedlings (10 seedlings per ecotype) were also used as control. Therefore, for this purpose, the two mentioned isolates were re-cultured on PDA medium and incubated under a 12-hr light/dark condition with a mixture of fluorescent light and near-ultraviolet at 25±3°C for 10 days. In addition, the seedlings’ stems were disinfested with 70% ethanol spraying before the inoculation program.
The seedlings were then inoculated by creating a surface wound in the stem with a scalpel at a 10-25 cm height from the collar. A 6-mm culture plug from actively growing hyphae was placed into the cut surface. Lastly, the inoculated part was wrapped with Parafilm (Ghaderi and Habibi, 2021). Similarly, a plug of sterile PDA medium was covered onto the wound in the control seedlings (as the control treatment). A piece of cotton soaked in sterile water was used to keep the inoculation point wet for 7 days (Linaldeddu et al., 2014). The control and inoculated seedlings were incubated at 25±3°C, a regime of 12 hr dark/light.
Water treatments and experimental design.
Water stress treatments were performed under greenhouse conditions after the inoculation of seedlings. Two irrigation regimes including control (non-stress) and intense stress (drought stress) were applied when 50±5%, and 90±5% of the total available soil moisture was depleted from the root-zone, respectively. Then, soil moisture was measured based on standard gravimetric methods at the root-zone (Carter and Gregorich, 2007). The irrigation intervals both during the experiment and between the two irrigation treatments were varied depending on soil moisture conditions. In order to perform water treatments, the pot experiment entailed three factors (moisture levels, ecotypes, and isolates) with three replications of four seedlings per treatment arranged in a randomized complete block design. The treatment combinations for each ecotype/region included: S0 I0 (non-stress and non-fungal inoculation), S0 I1 (non-stress and inoculation with Iran-G1), S0 I2 (non-stress and inoculation with M70), S1 I0 (water stress and non-fungal inoculation), S1 I1 (water stress and inoculation with G1), and S1 I2 (water stress and inoculation with Iran-M70). Therefore, each treatment combination had three replicates of eight seedlings per ecotype. The pot experiment lasted 12 weeks, from early June to early August 2019. The inoculated/non-inoculated seedlings under moisture environments were checked on a weekly basis for the induction of the symptoms such as leaf yellowing and early leaf browning for 12 weeks.
Measurements.
Following the 12-week period, the bark on the stem was removed and necrotic lesion length, as one of the initial symptoms of the development charcoal canker disease in inoculated tissue, was determined by using a Leaf Area Meter tool (Fanavaran Alborz Andisheh Company, Tehran, Iran). In order for the evaluation of the pathogenic capacity of the isolates used i.e., Iran-G1 and Iran-M70 in the seedlings were treated with two different irrigation regimes, the Koch's postulates were also used. For this purpose, by removing tissue section for the edge of the lesion at 3 to 5 mm intervals and the implantation onto PDA medium 25°C for colony growth, the re-isolation of fungi pathogens was carried out (Linaldeddu et al., 2014).
Statistical analysis.
All data for normality using the Kolmogorov-Smirnov test (Massey, 1951) were tested prior to being subjected to the combined analysis of variance (ANOVA) in line with the model of randomized complete block design using the GLM procedure of SAS statistical program (SAS Institute Inc., Cary, NC, USA). The least significant difference test (P≤0.05) was used to assess the significant variations between the means (pairwise comparisons).
Results
Identification of oak charcoal canker disease agent.
The initial symptoms of charcoal canker were almost nonspecific such as leaf yellowing, early leaf browning, and diminishing of canopy cover. Symptoms in advanced states of diseased indicated wilting, foliar rolling, production of carbonaceous stroma in the trunks and branches of the oak trees, dieback which eventually led to tree mortality (Fig. 1A-F).
In total, 120 isolates were obtained from declining oak trees in Kohgiluyeh & Boyer-Ahmad (60 isolates) and Fars provinces (60 isolates) in 2018-2019 (Supplementary Table 1), of which only one isolate was collected from each infected tree. All Biscogniauxia isolates were efficiently recovered from September to the end of November. Nevertheless, no Biscogniauxia isolate was obtained from tissues with symptoms and cankers from December until the end of February.
Morphological identification.
Based on morphological characters, isolates were recognized as Biscogniauxia mediterranea species. In general, the cultures showed colonies with white colony to gray with almost aerial mycelium on PDA medium and the back of colonies was dark-brown to black (Fig. 2A, B). The stromata were applanate, almost black surface, carbonaceous tissue. The perithecia were ovoid to tubular with black coarsely papillate ostioles. The asci were short stipitate with an amyloid ring. The ascospores were ellipsoid with narrowly rounded ends, with a spore-length germ slit only on one-side, brownish-black, lacking a hyaline sheath, 13-18×5.5-8 µm. Conidiogenesis holoblastic, Conidiogenous cell proliferation was essentially sympodial. The conidiophores were indistinguishable from vegetative hyphae; Conidia were subhyaline to hyaline, ovoid, ellipsoid, or subglobose, 9-12×3.5-5 mm (Fig. 2C-G). The maximum, minimum, and optimum temperatures for growth were 42°C, 10°C, and 30°C on PDA medium, respectively.
Fig. 2.
Colony growth of Biscogniauxia mediterranea on potato dextrose agar medium at 30°C (A); the reverse of the colonies (B); stromata exposed by dehiscence of overlying bark (C-E); ascospores, with a spore-length germ slit only on one-side (F); anamorph showing conidia and conidiophores (G). Scale bars=10 μm.

According to the results of ANOVA, significant differences (P<0.01) were observed among the isolates for the measured variable (growth rate) in the morphological assay (data not shown), indicating high levels of variation in the studied isolates. So that, the highest growth rate (11 cm) belonged to the Iran-G1 and Iran-M70 isolates from the Gachsaran and Mamasani regions, respectively (data not shown).
Phylogenetic analysis.
PCR amplification and sequencing were successful for all isolates. Obtained sequences of Biscogniauxia isolates were submitted in GenBank (Table 1).
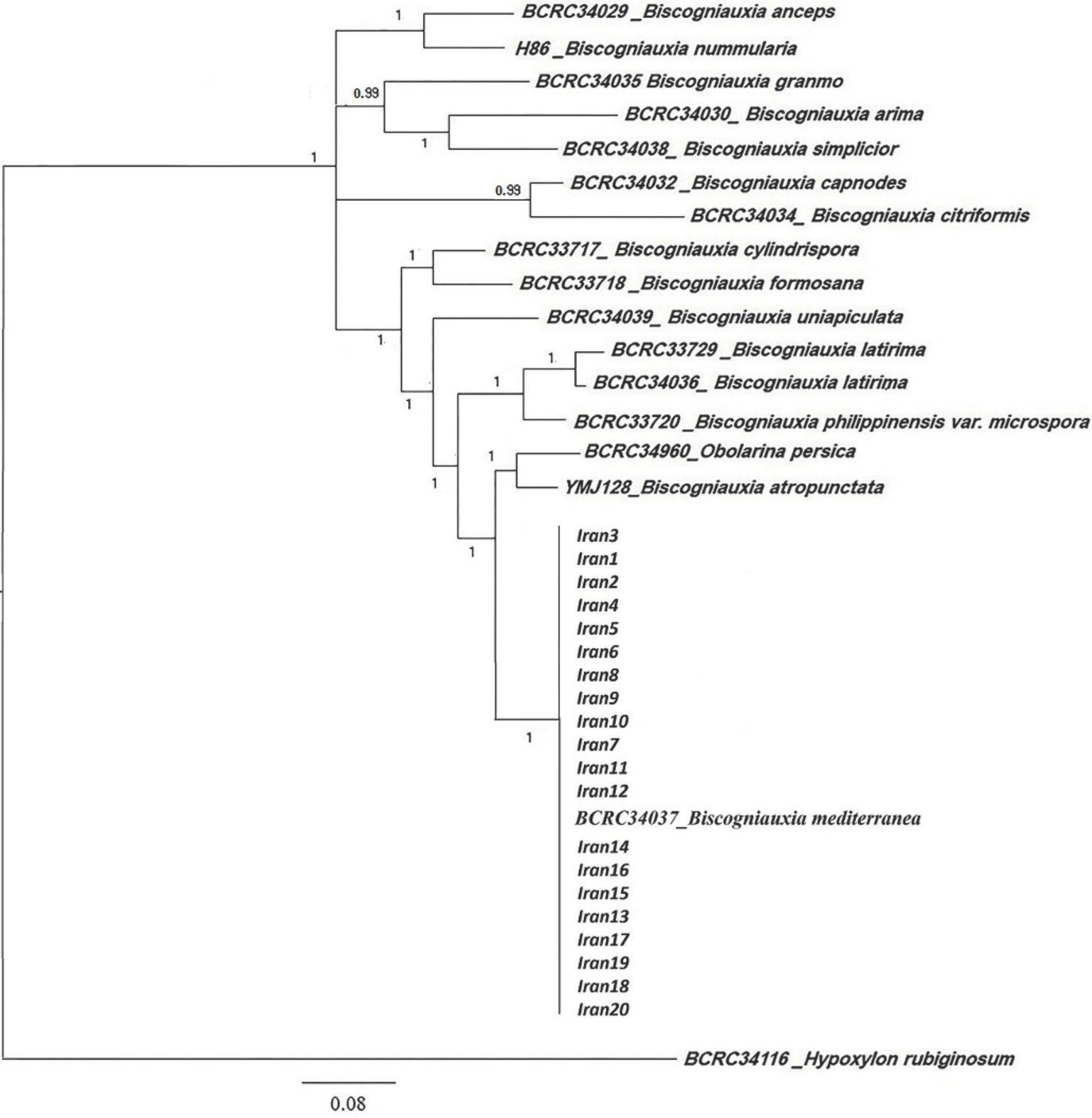
The aligned datasets for βtub, act, and ITS-rDNA consisted of 1,108, 332, and 846 characters, respectively. The aligned multigene datasets contained 2,276 characters, of which 691 were informative. An exhaustive list of aligned multigene dataset is available in Supplementary Table 1. The two phylogenetic analysis methods, including MP and Bayesian generated trees with similar topologies amongst species. The most appropriate model chosen by model test based on BIC was the general time reversible nucleotide substitution model with gamma-distributed rate variation and a proportion of invariable sites (GTR+I+G). The topology and branch lengths of the phylogenetic inferences are displayed in Fig. 3 and distinguished the phylogenetic position of isolates Iran 1 to Iran 20 using a Bayesian analysis of the combined data set of βtub, act, and ITS-rDNA sequences. The multigene phylogenies of combined dataset of βtub, act, and ITS-rDNA sequences revealed that isolates Iran1-Iran20 is a monophyletic group with B. mediterranea (strain: BCRC34037) in a well-supported clade (posterior probability=100).
Fig. 3.
Phylogram derived from Bayesian inference analysis of Btub, act, and internal transcribed spacer-rDNA dataset of Biscogniauxia. Bayesian posterior probabilities (%) are indicated above the branches. Hypoxylon rubiginosum is used as outgroup taxon. Scale bars=0.05 changes per site per branch.
Pathogenicity.
The effects of inoculation treatments (non-inoculation and inoculation) on a 2-year-old oak seedling were evaluated by pathogenicity test in greenhouse conditions. For this section, two isolates of Iran-G1 and Iran-M70 were selected, because the two mentioned isolates showed higher growth rate in the morphological evaluation. The results indicated that the identified species B. mediterranea was pathogenetic and the tested isolates caused brown to dark-brown lesions on the seedling's stem. The brown lesions spread up and down of the inoculation point and cross-section of the stem in infected areas showed wedge-shaped necrosis of vascular tissue (Fig. 4). So that the development of the necrosis in the inoculated stem was approximately 3-5 mm. In addition to stem tissue, in pathogenicity assay of B. mediterranea disease symptoms including yellowing and necrosis of leaves were appeared on inoculated seedlings 8 weeks after inoculation under greenhouse conditions (Fig. 4). In addition, the inoculated isolates were successfully re-iso-lated from brown lesions and Koch's postulates confirmed. Besides, no symptoms of disease were not detected on control seedlings (non-inoculation treatment) and no fungi pathogen was re-isolated.
Fig. 4.
Inoculation of two-years-old oak seedlings (Quercus brantii) with Biscogniauxia mediterranea after two months under greenhouse conditions: necrotic lesions area on 2-year-old oak seedlings without water stress (A), necrotic lesions area on 2-year-old oak seedlings with water stress (B), vascular necrosis in cross-section (C), and control treatment (no symptom) (D, E).

Pot-experiment and moisture treatments.
Effects of the moisture treatments (non-stress and intense drought stress) on the non-inoculated and inoculated seedlings (ecotypes) were evaluated in greenhouse conditions. The results of combined ANOVA indicated that the effects of moisture environment and isolate were highly significant for the trait of necrotic lesion length. Interactions of moisture environment×ecotype and moisture environment×isolate were also significant for the measured trait. These significant interactions revealed that the response of ecotypes and isolates varied in different environments for the measured character. However, the effects of ecotype, moisture environment×isolate×ecotype as well as isolate×ecotype were not significant (Supplementary Table 2). The non-sig-nificant effect of isolate×ecotype indicated that the ecotypes showed a similar response to the tested isolates (Supplementary Table 2).
Table 2.
Mean comparisons for the necrotic lesion length caused by the IS1 and IS2 in the five ecotypes of Quercus brantii under moisture environments
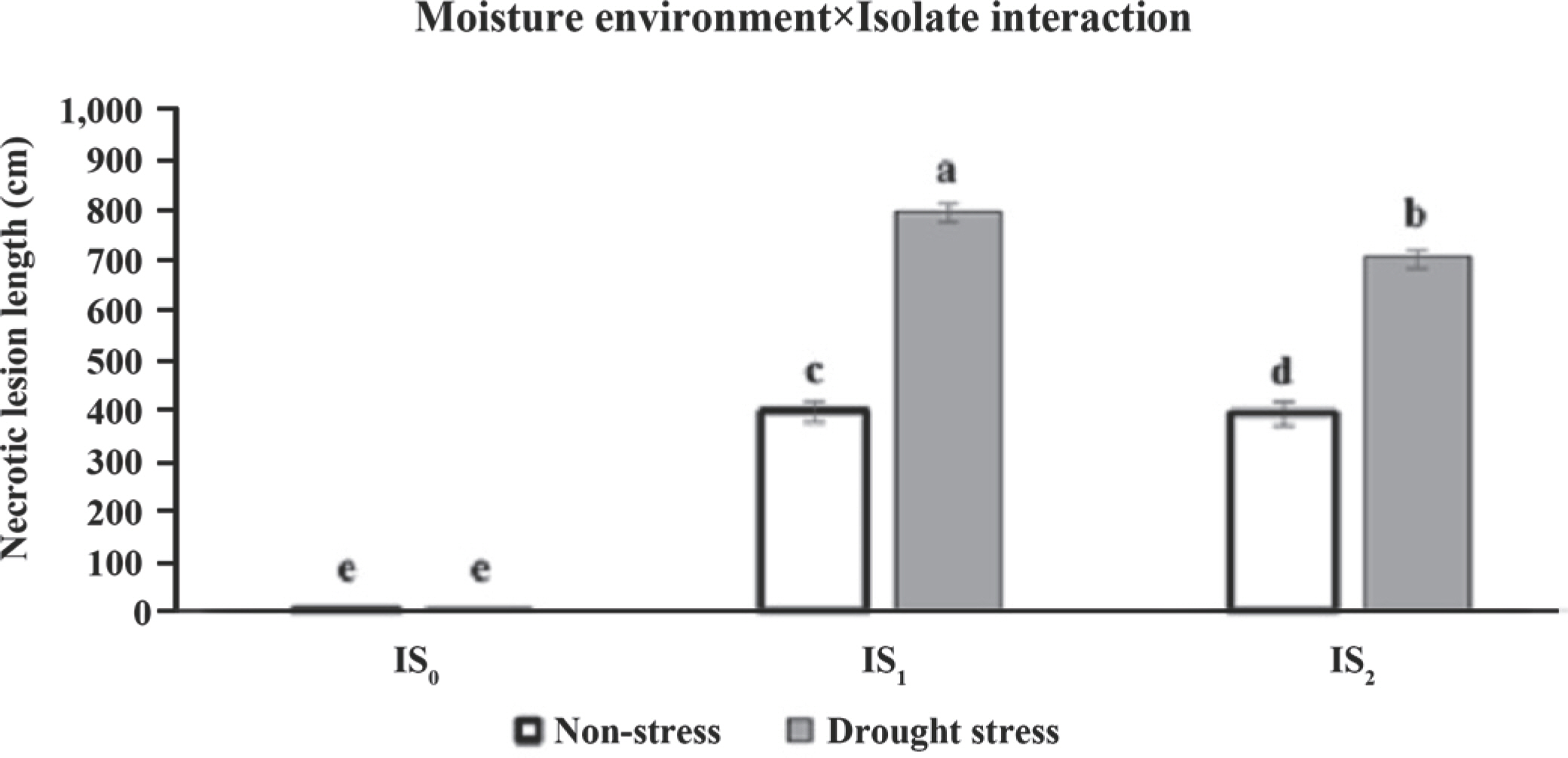
Regarding the moisture environment×isolate interaction, the comparison with the control moisture treatments (non-stress) revealed the necrotic lesion length in inoculated treatments by Iran-G1 (IS1) and Iran-M70 (IS2) increased (399.03 vs. 799.700 and 396.00 vs. 710.82 cm) under intense drought stresses. Meanwhile, under the same conditions, the length of IS1 induced-necrotic lesion was significantly wider than the IS2 in both moisture environments (Fig. 5).
Fig. 5.
Mean comparisons of necrotic lesion length in inoculation treatments (non-inoculation: IS0; inoculation by Iran I and Iran II iso-lates; IS1 and IS2, respectively) under two moisture environments (normal and intense stress) in the pot experiment. Data are pre-sented as mean±standard error of the mean. Different letters above the means reveal significant difference among the inoculation treatments according to the least significant difference test (P<0.05).
On the other hand, mean comparisons for the necrotic lesion length in the five ecotypes of Q. brantii under two moisture environments revealed that the response of ecotypes was varied in different environments (Table 2). Hence, under control conditions, the necrotic lesion length caused by the IS1 ranged from 398.22 cm (Dehdasht) to 400.31 cm (Basht), but there were no significant differences across the ecotypes (Table 2). However, the length of the necrotic lesion increased significantly when drought stress was applied. The length of IS2-induced-necrotic lesion was not extensive for ecotypes under control conditions, but expanded significantly, when the ecotypes were exposed to drought stress (Table 2). Meanwhile, there were no significant differences among the ecotypes in response to the tested isolates in each moisture environment (Table 2).
Discussion
In Iran, charcoal canker disease is notably a major problem in the southern regions of the Zagros forests. In this sur-vey, morphological and phylogenetic analyses of the DNA sequence data of βtub, act, and ITS-rDNA indicated that B. mediterranea is as the causal agent of charcoal canker on Q. brantii in southern Iran. Therefore, it seems that the present research conducted a comprehensive evaluation in southern Iran (two provinces/eight regions), in terms of molecular and morphological properties of causal agent of charcoal canker on Q. brantii, which is expected to provide new insights into the protection of oak forests in these areas. Generally, from early September to late March 2018-2019, surveys were conducted to identify charcoal canker disease in the oak forests of South Zagros, B. mediterranea single-spore isolates were obtained only from samples which were collected from September to November. In contrast, no isolates from the perithecial stromata were obtained in late autumn through mid-winter because the stromata tissues were rotten and easily crushed. Consequently, the ascospores of these samples were not able to germinate in the laboratory. Thus, our results were consistent with the previous findings of Safaee et al. (2017) which showed some stromatic samples did not contain live ascospores. Therefore, in field investigations, the status of stromatic specimens should be considered in terms of active or inactive for sampling purposes. We did not isolate Obolarina persica from the Persian oak trees with decline symptoms, while previous surveys have shown that this species is the other causal agent of charcoal canker in Persian oak trees in Ilam (Mirabolfathy et al., 2011), Kermanshah, Kurdestan and Lorestan (Safaee et al., 2016) provinces in western Iran.
B. mediterranea was pathogenic and caused dark-brown lesions in the inner bark on all 2-year-old oak seedlings from five different regions in Kohgiluyeh & Boyer-Ahmad prov-ince. Our results are in correspondence with pathogenicity test by Safaee et al. (2015) because they confirmed pathogenicity of B. mediterranea on branches and seedlings of Q. infectoria, Q. brantii, and Q. libani.
Based on the results obtained from the pot experiment (section II), drought stress increased lesion development in the stem of Q. brantii seedlings inoculated with B. mediterranea (100.41% and 79.48% increase for Iran-G1 and Iran-M70 isolates, respectively under drought conditions compared to non-stress conditions). This result indicated that drought stress and the occurrence of subsequent changes in the water content of plant tissues could affected the development of the studied fungal pathogen i.e., B. mediterranea (Vannini et al. 2009). In addition, a strong relationship between environmental stresses (drought) and the potential to cause pathogenesis, wood rot, canker, and dieback in different oak species was recorded by pathogens such as B. mediterranea (Capretti and Battisti 2007; Corcobado et al. 2014; Ghanbary et al. 2017; Linaldeddu et al. 2011; Vannini et al. 2009). The results obtained from moisture environment×isolate interaction indicated that drought could lead to further development and acceleration of negative effects of the charcoal canker disease on the host plant.
Drought has substantial influences on the biochemical, metabolical, morphological, and physiological aspects of plants (Faraji et al., 2020; Goharrizi et al., 2020). Reduction of plant water potential in drought conditions is one of the common and primary physiological reactions to osmotic regulation and plant water conservation to increase drought resistance (Ranjbarfordoei et al., 2013) that has been studied in different species of oak (Aranda et al., 2005; Capretti and Battisti, 2007; Cotrozzi et al., 2016) and other plants/trees (Goharrizi et al., 2020; Saeidnia et al., 2018). In addition, Chanbary et al. (2017) reported a direct correlation between the pathogenicity of the charcoal canker disease and oak leaf water potential.
On the other hand, the effect of reducing water potential in host tissues as an initiator of transition from the latent phase to the pathogenic stage in B. mediterranea on Quercus species has been reported (Linaldeddu et al., 2011; Vannini et al., 2009). This change in the phase of pathogen i.e., the transition from the latent to aggressive due to drought and reduced water potential, weakens plant's defense systems against the spread of pathogenic fungi (Linaldeddu et al., 2011). Indeed, consequences of drought stress, manifested as declined relative water content could be associated with a decrease in the activity of antioxidative defense enzymes (Goharrizi et al., 2020; Saeidnia et al., 2018). On the other hand, it seems that the studied fungus colonizes most of the xylem's tissue under drought conditions. Furthermore, according to producing cankers on vascular tissues of infected seedlings by B. mediterranea, it seems that the mentioned tissues could affect the water status of plant including changes in water flow to the leaves. The role of fungal pathogens in the reduction of the water potential of woody vessels has been reported (Capretti and Battisti, 2007).
In the present study, what was also investigated was whether the exposure of inoculated oak seedlings by pathogen of B. mediterranea to water deficient stress could affect the spread/severity of charcoal canker disease. Our results support previous findings across many plant species that drought stress is destructive and could lead to further development and acceleration of negative effects of charcoal canker disease on Q. brantii in southern Iran.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Supplement1
Supplement1 Print
Print






