

ņä£ļĪĀ
ļ┐īļ”¼Ēś╣ņäĀņČ®(Meloidogyne spp.)ņØĆ ņŗØļ¼╝ ļ┐īļ”¼ņŚÉ ņĀłļīĆĻĖ░ņāØĒĢśļ®░ ĒåĀļ¦łĒåĀ, Ļ│ĀĻĄ¼ļ¦ł, ņ░ĖņÖĖ, ņśżņØ┤ ļō▒ ņŻ╝ņÜö ļåŹņ×æļ¼╝ņŚÉ ņŗ¼Ļ░üĒĢ£ Ēö╝ĒĢ┤ļź╝ ņĢ╝ĻĖ░ĒĢśļŖö ĒåĀņ¢æ ļ│æņøÉĻĘĀ ņżæņØś ĒĢśļéśņØ┤ļŗż. ņĀä ņäĖĻ│äņĀüņ£╝ļĪ£ ņ×æļ¼╝ Ēö╝ĒĢ┤ņØś ņĢĮ 11%Ļ░Ć ņäĀņČ®ņŚÉ ņØśĒĢ£ Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ņ£╝ļ®░(McCart, 2009), ĻĄŁļé┤ņØś Ļ▓ĮņÜ░ņŚÉļÅä Ļ▓ĮļČü ņä▒ņŻ╝, ņČ®ļé© Ļ│ĄņŻ╝ ļō▒ņØś ĒżņןņŚÉņä£ ļ┐īļ”¼Ēś╣ņäĀņČ®ņŚÉ ņØśĒĢ£ ļ│æ ļ░£ņāØ Ēö╝ĒĢ┤ņé¼ļĪĆĻ░Ć ļ│┤Ļ│ĀļÉśĻ│Ā ņ׳ļŗż(Park ļō▒, 2011, 2012). Ēśäņ×¼Ļ╣īņ¦Ć ļÅÖņĀĢļÉ£ ņĢĮ 90ņŚ¼ ņóģ ņØ┤ņāüņØś ļŗżņ¢æĒĢ£ ļ┐īļ”¼Ēś╣ņäĀņČ® ņżæņŚÉņä£ļÅä M. incognita, M. arenaria, M. javanica, M. hapla ļō▒ņØĆ ļäōņØĆ ĻĖ░ņŻ╝ ļ▓öņ£äļź╝ Ļ░Ćņ¦Ćļ®░ ņ×æļ¼╝ ņłśĒÖĢļ¤ēņØä Ļ░Éņåīņŗ£ĒéżĻĖ░ ļĢīļ¼ĖņŚÉ Ļ▓ĮņĀ£ņĀüņ£╝ļĪ£ ņżæņÜöĒĢ£ ļ┐īļ”¼Ēś╣ņäĀņČ®ņ£╝ļĪ£ Ļ░äņŻ╝ĒĢśĻ│Ā ņ׳ļŗż(Kim ļō▒, 2011, 2014; TrudgillĻ│╝ Blok, 2001). ļ┐īļ”¼Ēś╣ņäĀņČ®ņŚÉ ņØśĒĢ┤ Ļ░ÉņŚ╝ļÉ£ ņŗØļ¼╝ņ▓┤ņØś ļ┐īļ”¼ņŚÉļŖö Ļ▒░ļīĆĒĢ£ Ēś╣ņØ┤ ĒśĢņä▒ļÉśļ®░, ņØ┤ļĪ£ ņØĖĒĢ┤ ņŗØļ¼╝ņ▓┤ļŖö ļ¼╝Ļ│╝ ņśüņ¢æļČäņØś ĒØĪņłśļéś ļ»Ėļäżļ×äņØś ņØ┤ļÅÖ ļō▒ņŚÉ ļ░®ĒĢ┤ļź╝ ļ░øĻ▓ī ļÉśņ¢┤ ņĀĢņāüņĀüņØĖ ņāØņןņØ┤ ņ¢┤ļĀżņøīņ¦äļŗż(WilliamsonĻ│╝ Hussey, 1996). ņØ┤ņ▓śļ¤╝ Ēś╣ņØ┤ ĒśĢņä▒ļÉ£ ņŗØļ¼╝ļ┐īļ”¼ņØś Ēæ£ļ®┤, ļśÉļŖö ĻĘĖ Ēś╣ņØś ļé┤ļČĆņŚÉļŖö ņĢīņØ┤ ņĪ┤ņ×¼ĒĢśĻ▓ī ļÉśļ®░, ņĀüņĀłĒĢ£ ņĪ░Ļ▒┤ņØ┤ ļ¦īņĪ▒ļÉśļ®┤ ņĢīņŚÉņä£ J2 ļŗ©Ļ│äņØś ņ£ĀņČ®ņØ┤ ļČĆĒÖöļÉ£ļŗż(PerryņÖĆ Moens, 2006). J2 ņ£ĀņČ® ļŗ©Ļ│äņØś ļ┐īļ”¼Ēś╣ņäĀņČ®ņØĆ ņŗØļ¼╝ņ▓┤ņØś ļ┐īļ”¼ ļüØņ£╝ļĪ£ ņ╣©Ēł¼ĒĢśņŚ¼ ņäĖĒż ņé¼ņØ┤ļź╝ ļÅīņĢäļŗżļŗłļŗżĻ░Ć ņ£ĀĻ┤ĆņåŹņŻ╝ļĪ£ ņ╣©ņ×ģĒĢśņŚ¼ ļ┐īļ”¼Ēś╣ņØä ņ×¼ĒśĢņä▒ĒĢśĻ▓ī ļÉ£ļŗż(Caillaud ļō▒, 2008).
ņŗØļ¼╝ĻĖ░ņāØņäĀņČ®ņŚÉ ņØśĒĢ£ Ēö╝ĒĢ┤ļź╝ Ļ░Éņåīņŗ£ĒéżĻĖ░ ņ£äĒĢ┤ ņ£żņ×æ, Ļ░ØĒåĀ, Ēā£ņ¢æņŚ┤ ņåīļÅģ, ļŗ┤ņłśņ▓śļ”¼ ļō▒Ļ│╝ Ļ░ÖņØĆ ļ¼╝ļ”¼ņĀü ļ░®ņĀ£ļź╝ ļ╣äļĪ»ĒĢśņŚ¼ ņĀĆĒĢŁņä▒ ņŗØļ¼╝ ĒÆłņóģ ņ×¼ļ░░, ĒÖöĒĢÖņĀü ļ░®ņĀ£ļ▓Ģ ļō▒ņØ┤ Ļ│ĀļĀżļÉ£ļŗż(Park ļō▒, 2012; PerryņÖĆ Moens, 2006). ļ┐īļ”¼Ēś╣ņäĀņČ®ļ│æņØś ĒÖöĒĢÖņĀü ļ░®ņĀ£ ņłśļŗ©ņ£╝ļĪ£ļŖö ļ®öĒŗĖ ļĖīļĪ£ļ¦łņØ┤ļō£(methyl bromide) ļō▒ņØś Ēøłņ”ØņĀ£ņÖĆ ĒżņŖżĒŗ░ņĢäņĀ£ņØ┤ĒŖĖ(fosthiazate) ļō▒ņØś ņé┤ņäĀņČ®ņĀ£Ļ░Ć ņŻ╝ļĪ£ ņØ┤ņÜ®ļÉśņ¢┤ ņÖöļŗż. ĒĢśņ¦Ćļ¦ī ņØ┤ļ¤¼ĒĢ£ ĒÖöĒĢÖņĀü ļ░®ņĀ£ļŖö ņØĖņČĢņŚÉ ļīĆĒĢ£ ņ£äĒŚśņä▒Ļ│╝ ņ×öļźśļÅģņä▒, ĒÖśĻ▓ĮņśżņŚ╝, ņāØĒā£Ļ│ä ĻĄÉļ×Ć ļō▒Ļ│╝ Ļ░ÖņØĆ ļŗżņ¢æĒĢ£ ļ¼ĖņĀ£ņĀÉņØä ļģĖņČ£ĒĢśĻ│Ā ņ׳ņ£╝ļ®░, ņØ┤ļĪ£ ņØĖĒĢ┤ ļ│┤ļŗż ņĢłņĀäĒĢ£ ņé┤ņäĀņČ®ņĀ£ņØś Ļ░£ļ░£ņØ┤ ņÜöĻĄ¼ļÉśĻ│Ā ņ׳ļŗż. ĒÖśĻ▓Į ņ╣£ĒÖöņĀüņ£╝ļĪ£ ņŗØļ¼╝ĻĖ░ņāØņäĀņČ®ņØä ļ░®ņĀ£ĒĢśĻĖ░ ņ£äĒĢśņŚ¼ ņŗØļ¼╝ ņČöņČ£ļ¼╝ņØ┤ļéś ļ»ĖņāØļ¼╝, ļśÉļŖö ļ»ĖņāØļ¼╝ņØś ņØ┤ņ░©ļīĆņé¼ņé░ļ¼╝ņØä ņØ┤ņÜ®ĒĢ£ ļ░®ļ▓ĢļōżņØ┤ Ļ│ĀļĀżļÉśĻ│Ā ņ׳ņ£╝ļ®░, ļÅÖņŗ£ņŚÉ ļ¦ÄņØĆ ņŚ░ĻĄ¼ļōżņØ┤ ņ¦äĒ¢ēļÉśņ¢┤ ņśżĻ│Ā ņ׳ļŗż(AkhtarņÖĆ Malik, 2000; Kerry, 2000; Oka ļō▒, 2000; Rodriguez-Kabana ļō▒, 1987; Sharon ļō▒, 2001). Pseudomonas fluorescensņØś ļ░░ņ¢æņŚ¼ņĢĪņØĆ in vitroņāüņŚÉņä£ ņäĀņČ®ņØś ņĢī ļČĆĒÖöņ£© Ļ░Éņåī ļ░Å M. javanicaņØś ņ£ĀņČ®ņŚÉ ļīĆĒĢ£ ņé┤ņäĀņČ® ĒÜ©Ļ│╝ļź╝ ļ│┤ņśĆņ£╝ļ®░, ņØ┤ļŖö P. fluorescensņØś ņØ┤ņ░©ļīĆņé¼ņé░ļ¼╝ņØĖ 2,4-diacetylphloroglucinolņŚÉ ņØśĒĢ£ ĒÜ©Ļ│╝ļĪ£ ļ░ØĒśĆņĪīļŗż(SiddiquiņÖĆ Shaukat, 2003). ļśÉĒĢ£ Trichoderma harzianum BIĻ│╝ 27ņóģņØś ņŗØļ¼╝ņ▓┤ļĪ£ļČĆĒä░ ņ¢╗ņØĆ ļŗżņ¢æĒĢ£ essential oilļÅä M. javanicaņØś ņ╣śņé¼ņ£©ņŚÉ ņśüĒ¢źņØä ņżĆļŗżļŖö ņŚ░ĻĄ¼ Ļ▓░Ļ│╝Ļ░Ć ņ׳ļŗż(Oka ļō▒, 2000; SahebaniņÖĆ Hadavi, 2008). ņĄ£ĻĘ╝ Jang ļō▒(2016)ņØĆ Aspergillus niger F22ņØś ļ░░ņ¢æņŚ¼ņĢĪņØ┤ M. incognita J2 ņ£ĀņČ®ņØś ņ╣śņé¼ņ£© ņ”ØĻ░Ć ļ░Å ņäĀņČ® ņĢīņØś ļČĆĒÖöņ£© Ļ░Éņåī Ļ▓░Ļ│╝ļź╝ ņ┤łļלĒĢ£ļŗżĻ│Ā ļ│┤Ļ│ĀĒĢśņśĆņ£╝ļ®░, ņØ┤ļŖö A. niger F22 ĻĘĀņŻ╝Ļ░Ć ņāØņé░ĒĢśļŖö oxalic acidļĪ£ļČĆĒä░ ĻĖ░ņØĖĒĢ£ ĒÜ©Ļ│╝ļØ╝Ļ│Ā ļ│┤Ļ│ĀĒĢśņśĆļŗż.
ĒåĀņ¢æņŚÉ ņä£ņŗØĒĢśļŖö ņäĀņČ®ņØĆ ļŗżļźĖ ĻĘĀļźśļōżņŚÉ ņØśĒĢ┤ ņ×ÉĻĘ╣ ļ░øņØä ņłś ņ׳ņ¢┤ ĒåĀņ¢æļ»ĖņāØļ¼╝ņØä ĒÖ£ņÜ®ĒĢ£ ņāØļ¼╝ĒĢÖņĀü ņäĀņČ®ļ░®ņĀ£ļ▓ĢņØ┤ ļ¦ÄņØĆ Ļ┤Ćņŗ¼ņØä ļ░øņĢäņśżĻ│Ā ņ׳ļŗż. ļ░®ņäĀĻĘĀņØĆ ĒåĀņ¢æņŚÉ ņä£ņŗØĒĢśļŖö ņäĖĻĘĀņ£╝ļĪ£, ĻĘĀņé¼ņØś ĒśĢĒā£ļĪ£ ņ”ØņŗØĒĢśļ®┤ņä£ ļŗżņ¢æĒĢ£ ņØ┤ņ░©ļīĆņé¼ņé░ļ¼╝ņØä ņāØņä▒ĒĢ£ļŗż(SiddiquiņÖĆ Mahmood, 1999). Ēśäņ×¼Ļ╣īņ¦Ć ņĢīļĀżņ¦ä ņ▓£ņŚ░ ĒĢŁņāØļ¼╝ņ¦łņØś 60%-70%Ļ░Ć ļ░®ņäĀĻĘĀņ£╝ļĪ£ļČĆĒä░ ļ░£Ļ▓¼ļÉśņŚłņ£╝ļ®░, 1970ļģäļīĆņŚÉ ĻĘĖ ņłśĻ░Ć ņĄ£ļīĆņ╣śļź╝ ņ░ŹņØĆ ņØ┤ĒøäļĪ£ļÅä ļ¦żļģä ņØ┤ļōżņØ┤ ļé┤ļŖö ĒĢŁņāØļ¼╝ņ¦łņØ┤ ĻŠĖņżĆĒ׳ ļ│┤Ļ│ĀļÉśĻ│Ā ņ׳ļŗż(B├®rdy, 2005; Watve ļō▒, 2001). Avermectin, bafilomycin, fervenulin, 6ŌĆÖ-methyl-fungichromin ļō▒ņØ┤ ļ░®ņäĀĻĘĀņŚÉ ņØśĒĢ┤ ņāØņé░ļÉśļŖö ņé┤ņäĀņČ® ļ¼╝ņ¦łļĪ£ ņĢīļĀżņĀĖ ņ׳ņ£╝ļ®░, ņØ┤ļōż ļ¼╝ņ¦łļōżņØĆ ņé┤ņäĀņČ® ĒÖ£ņä▒ ņØ┤ņÖĖņŚÉļÅä ļ│æņøÉņä▒ Ļ│░ĒīĪņØ┤ļéś ļ░ĢĒģīļ”¼ņĢäņŚÉ ļīĆĒĢ£ ĒĢŁĻĘĀ ĒÖ£ņä▒ņØä Ļ░¢ļŖö Ļ▓āņ£╝ļĪ£ ļ│┤Ļ│ĀļÉśņ¢┤ ņ׳ļŗż(Lacey ļō▒, 1995; Putter ļō▒, 1981; Ruanpanun ļō▒, 2011; Zeng ļō▒, 2013). ĒĢśņ¦Ćļ¦ī ņØ┤ļ¤¼ĒĢ£ ļŗżņ¢æĒĢ£ ņŚ░ĻĄ¼Ļ▓░Ļ│╝ņŚÉļÅä ļČłĻĄ¼ĒĢśĻ│Ā ĻĖĖĒĢŁ ļ░®ņäĀĻĘĀņØä ņ¦üņĀæ ņØ┤ņÜ®ĒĢ£ ļ»ĖņāØļ¼╝ ļåŹņĢĮ Ļ░£ļ░£ņØĆ ļ»Ėļ╣äĒĢ£ ņŗżņĀĢņØ┤ļŗż.
ļö░ļØ╝ņä£ ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö ĒåĀņ¢æņŚÉņä£ ļČäļ”¼ĒĢ£ 670ņŚ¼ Ļ░£ņØś ļ░®ņäĀĻĘĀņØä ļīĆņāüņ£╝ļĪ£ ļ┐īļ”¼Ēś╣ņäĀņČ®(M. incognita)ņŚÉ ļīĆĒĢ£ ņé┤ņäĀņČ® ĒÖ£ņä▒ ņĪ░ņé¼ļź╝ ņŗżņŗ£ĒĢśņśĆĻ│Ā, ņØ┤ ņżæ Ļ░Ćņן ņÜ░ņłśĒĢ£ ĒÖ£ņä▒ņØä Ļ░¢ļŖö Streptomyces flavogriseus KRA15-528 ĻĘĀņŻ╝ļź╝ ņäĀļ░£ĒĢśņśĆļŗż. ņäĀļ░£ļÉ£ ĻĘĀņŻ╝ņŚÉ ļīĆĒĢ£ in vivoņÖĆ in vitro ņé┤ņäĀņČ® ĒÖ£ņä▒ ņĪ░ņé¼ļź╝ ĒåĄĒĢ┤ ņ╣£ĒÖśĻ▓Į ņé┤ņäĀņČ®ņĀ£ļĪ£ņä£ņØś Ļ░£ļ░£ Ļ░ĆļŖźņä▒ņØä Ļ▓ĆĒåĀĒĢśņśĆļŗż.
ņ×¼ļŻī ļ░Å ļ░®ļ▓Ģ
ļ░®ņäĀĻĘĀņØś ļ░░ņ¢æ ļ░Å ļ░░ņ¦ĆņĪ░Ļ▒┤
2015ļģäļÅäņŚÉ ļŗżņ¢æĒĢ£ ņ¦ĆņŚŁņØś ĒåĀņ¢æņ£╝ļĪ£ļČĆĒä░ ļ░®ņäĀĻĘĀņØä ļČäļ”¼ĒĢśņśĆņ£╝ļ®░, ļČäļ”¼ļÉ£ ļ░®ņäĀĻĘĀņØś ņé┤ņäĀņČ® ĒÖ£ņä▒ņØä ņĪ░ņé¼ĒĢśĻ│Āņ×É ļŗżņØīĻ│╝ Ļ░ÖņØĆ ļ░░ņ¦Ć ļ░Å ļ░░ņ¢æņĪ░Ļ▒┤ņØä ņØ┤ņÜ®ĒĢśņśĆļŗż. ļ░®ņäĀĻĘĀņØĆ BennettŌĆÖs ĒĢ£ņ▓£ļ░░ņ¦Ć(1% glucose, 0.1% yeast extract, 0.2% Bacto-peptone, 0.1% beef extract, 2% agar) (Hesseltine ļō▒, 1954)ņŚÉ ļÅäļ¦ÉĒĢśņŚ¼ Ļ│Āņ▓┤ ļ░░ņ¢æĒĢ£ Ēøä, ĒśĢņä▒ļÉ£ ļŗ©ņØ╝ ņĮ£ļĪ£ļŗłļź╝ tryptic soy broth ļ░░ņ¦ĆņŚÉ ņĀæņóģĒĢśĻ│Ā 28┬░C, 150 rpmņŚÉņä£ 2ņØ╝ ļÅÖņĢł ņĀäļ░░ņ¢æĒĢśņśĆļŗż. ņĀäļ░░ņ¢æĒĢ£ ļ░®ņäĀĻĘĀņØĆ glucose-soybean meal-starch (GSS) ļ░░ņ¦Ć(1% soluble starch, 2% glucose, 2.5% soybean meal, 0.1% beef extract, 0.4% yeast extract, 0.2% NaCl, 0.025% K2HPO4, 0.1% CaCO3) (Kim ļō▒, 1989)ņŚÉ 1% (v/v)Ļ░Ć ļÉśļÅäļĪØ ņĀæņóģĒĢśĻ│Ā 28┬░C, 150 rpmņŚÉņä£ 7ņØ╝ ļÅÖņĢł ņ¦äĒāĢ ļ░░ņ¢æĒĢśņŚ¼ ņé┤ņäĀņČ® ĒÖ£ņä▒ ņĪ░ņé¼ņŚÉ ņé¼ņÜ®ĒĢśņśĆļŗż.
ļ┐īļ”¼Ēś╣ņäĀņČ®ņØś ņ”ØņŗØ ļ░Å ļČäļ”¼
Hwang ļō▒(2014)ņŚÉ ņØśĒĢ┤ ļČäļ”¼, ļÅÖņĀĢļÉ£ ļ┐īļ”¼Ēś╣ņäĀņČ®ņØĆ ņś©ņŗż(25┬░C-5┬░C)ņŚÉņä£ ŌĆśņä£Ļ┤æŌĆģŌĆÖ ĒÆłņóģņØś ĒåĀļ¦łĒåĀņŚÉ ņ”ØņŗØņŗ£ņ╝£ ņŗżĒŚśņŚÉ ņé¼ņÜ®ĒĢśņśĆļŗż. ņøÉņśłņÜ® ņāüĒåĀ(High; Punong, Gyeongju, Korea)ļź╝ ņ¦üĻ▓Į 5 cmņØś ĒöīļØ╝ņŖżĒŗ▒ ņ╗ĄņŚÉ ļäŻĻ│Ā ĒåĀļ¦łĒåĀļź╝ ĒīīņóģĒĢśņśĆļŗż. 3ņŻ╝Ļ░ä ņ×¼ļ░░ĒĢ£ ņ£Āļ¼śļź╝ ĒöīļØ╝ņŖżĒŗ▒ ĒżĒŖĖņŚÉ ņØ┤ņŗØĒĢ£ ļÆż ļ┐īļ”¼Ēś╣ņäĀņČ®ņØś ņĢīņØä ĒåĀļ¦łĒåĀ ĒĢ£ ņŻ╝ļŗ╣ 5,000Ļ░£ņö® ņĀæņóģĒĢśņśĆņ£╝ļ®░, 6ņŻ╝ ļÅÖņĢł ņ×¼ļ░░ĒĢśņŚ¼ Ēś╣ņØ┤ ņŗ¼ĒĢśĻ▓ī ĒśĢņä▒ļÉ£ ĒåĀļ¦łĒåĀ ļ┐īļ”¼ļź╝ ņ▒äņĘ©ĒĢśņśĆļŗż.
ļ┐īļ”¼ņŚÉ ņāØĻĖ┤ Ēś╣Ļ│╝ ļé£ļéŁņ£╝ļĪ£ļČĆĒä░ ļ┐īļ”¼Ēś╣ņäĀņČ® ņ£ĀņČ®(J2)ņØä ļČäļ”¼ĒĢśĻĖ░ ņ£äĒĢ┤ Ēś╣ņØ┤ ĒśĢņä▒ļÉ£ ĒåĀļ¦łĒåĀ ļ┐īļ”¼ļź╝ ļ¼╝ļĪ£ ņäĖņ▓ÖĒĢ£ Ēøä 1 cm Ēü¼ĻĖ░ļĪ£ ņĀłļŗ©ĒĢśņśĆļŗż. ĒåĀļ¦łĒåĀ ļ┐īļ”¼Ļ░Ć 0.5% ņ░©ņĢä ņŚ╝ņåīņé░ļéśĒŖĖļź©(sodium hypochlorite) ņÜ®ņĢĪņŚÉ ņ×ĀĻĖĖ ņĀĢļÅäļĪ£ ļäŻņØĆ Ēøä, ļČäņćäĻĖ░ļĪ£ 60ņ┤łĻ░ä ļČäņćäĒĢśņśĆļŗż. ļČäņćäĒĢ£ ņŗ£ļŻīļŖö 230 mesh ņ▓┤ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņ░īĻ║╝ĻĖ░ļź╝ Ļ▒░ļź┤Ļ│Ā 550 mesh ņ▓┤ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņĢīņØä ņłśņ¦æĒĢśņśĆļŗż(Barker ļō▒, 1985). ņ▓┤ņŚÉ ļ¬©ņØĖ ņäĀņČ® ņĢīņØĆ ļ®ĖĻĘĀņłśļĪ£ 3ļ▓ł ņäĖņ▓ÖĒĢ£ ļÆż ļČĆĒÖö ņ¢ĄņĀ£ ĒÖ£ņä▒ ņĪ░ņé¼ņŚÉ ņØ┤ņÜ®ļÉśĻ▒░ļéś, Baermann funnel ļ░®ļ▓ĢņØä ņØ┤ņÜ®ĒĢśņŚ¼ ņäĀņČ® ņĢīļĪ£ļČĆĒä░ ņ£ĀņČ®ņØä ļČäļ”¼ĒĢśņśĆļŗż(Southey, 1986).
ļ░®ņäĀĻĘĀ ļ░░ņ¢æņŚ¼ņĢĪņØ┤ ļ┐īļ”¼Ēś╣ņäĀņČ® ņ£ĀņČ®Ļ│╝ ņĢīņŚÉ ļ»Ėņ╣śļŖö ņśüĒ¢ź
GSS ļ░░ņ¦ĆņŚÉņä£ 7ņØ╝ ļÅÖņĢł ļ░░ņ¢æĒĢ£ ļ░®ņäĀĻĘĀ ļ░░ņ¢æņĢĪņØä 4┬░C, 6,500g ņĪ░Ļ▒┤ņŚÉņä£ 10ļČä ļÅÖņĢł ņøÉņŗ¼ļČäļ”¼ĒĢśņŚ¼ ņ¢╗ņ¢┤ņ¦ä ņāüļō▒ņĢĪņØä ņé¼ņÜ®ĒĢśņŚ¼ ņ£ĀņČ®ņŚÉ ļīĆĒĢ£ ņé┤ņäĀņČ® ĒÖ£ņä▒Ļ│╝ ņĢī ļČĆĒÖö ņ¢ĄņĀ£ ĒÖ£ņä▒ ņĪ░ņé¼ļź╝ ĒĢśņśĆļŗż. ļ©╝ņĀĆ, ņ£ĀņČ®ņŚÉ ļīĆĒĢ£ ņé┤ņäĀņČ® ĒÖ£ņä▒ņØä ņĪ░ņé¼ĒĢśĻĖ░ ņ£äĒĢ┤ ņäĀņČ® ĒśäĒāüņĢĪņØä 96-well tissue culture plate (Becton Dickinson, Franklin Lakes, NJ, USA)ņØś Ļ░ü wellņŚÉ ļäŻņØĆ ļŗżņØī ļ░®ņäĀĻĘĀ ļ░░ņ¢æņŚ¼ņĢĪņØś ļåŹļÅäĻ░Ć 10%, 5%, 2.5%, 1.25%Ļ░Ć ļÉśļÅäļĪØ ņ▓śļ”¼ĒĢśĻ│Ā wellļŗ╣ ņĄ£ņóģ ļČĆĒö╝Ļ░Ć 100 ╬╝l (ņĢĮ 50ļ¦łļ”¼ J2)Ļ░Ć ļÉśļÅäļĪØ ĒĢśņśĆļŗż. ņŗ£ļŻīĻ░Ć ĻĘĀņØ╝ĒĢśĻ▓ī ņä×ņØ┤ļÅäļĪØ 30ņ┤ł ļÅÖņĢł ĻĄÉļ░śĒĢśņśĆņ£╝ļ®░, ņäĀņČ®ņØś ņé░ņåīĻ▓░ĒĢŹĻ│╝ Ļ▒┤ņĪ░ļź╝ ļ░®ņ¦ĆĒĢśĻĖ░ ņ£äĒĢśņŚ¼ ņāüļīĆņŖĄļÅä 100%ņØś ĒöīļØ╝ņŖżĒŗ▒ ĒåĄņŚÉ ļäŻņØĆ Ēøä 25┬░CņŚÉņä£ ļ░░ņ¢æĒĢśņśĆļŗż. ņŗżĒŚśņØĆ 3ļ░śļ│ĄņØä ņłśĒ¢ēĒĢśņśĆņ£╝ļ®░, ņŗ£ļŻīņ▓śļ”¼ 48ņŗ£Ļ░ä, 72ņŗ£Ļ░ä, 96ņŗ£Ļ░ä Ēøä Ļ┤æĒĢÖĒśäļ»ĖĻ▓Į(CKX41; Olympus, Tokyo, Japan)ĒĢśņŚÉņä£ Ļ┤Ćņ░░ĒĢśņŚ¼ ņ¦üņäĀņ£╝ļĪ£ ļ╗Śņ¢┤ ņøĆņ¦üņ×äņØ┤ ņŚåļŖö ņäĀņČ®ņØä ņŻĮņØĆ ņäĀņČ®ņ£╝ļĪ£, ņøĆņ¦üņ×äņØ┤ ņ׳ļŖö ņäĀņČ®ņØä ņé┤ņĢäņ׳ļŖö ņäĀņČ®ņ£╝ļĪ£ ĒīÉļŗ©ĒĢśņŚ¼ ņé┤ņäĀņČ®ļźĀņØä ņĪ░ņé¼ĒĢśņśĆļŗż(Abbott, 1925). ņĢī ļČĆĒÖö ņ¢ĄņĀ£ ĒÖ£ņä▒ņØä ņĪ░ņé¼ĒĢśĻĖ░ ņ£äĒĢ┤ ņ£ĀņČ®ņŚÉ ļīĆĒĢ£ ņé┤ņäĀņČ® ĒÖ£ņä▒ņØä ņĪ░ņé¼ĒĢśĻĖ░ ņ£äĒĢ£ ļ░®ļ▓ĢĻ│╝ ļÅÖņØ╝ĒĢ£ ļ░®ļ▓ĢņØä ņé¼ņÜ®ĒĢśņśĆļŗż. ņäĀņČ® ņĢīņØä Ļ░ü wellņŚÉ ņĢĮ 50Ļ░£ņö® ļäŻņØĆ ļŗżņØī ļ░®ņäĀĻĘĀ ļ░░ņ¢æņŚ¼ņĢĪņØś ļåŹļÅäĻ░Ć 10%, 5%, 2.5%ļĪ£ ļÉśļÅäļĪØ ņ▓śļ”¼ĒĢśĻ│Ā wellļŗ╣ ņĄ£ņóģ ļČĆĒö╝Ļ░Ć 100 ╬╝lĻ░Ć ļÉśļÅäļĪØ ĒĢśņśĆļŗż. ļ░░ņ¢æņĪ░Ļ▒┤ņØĆ ņ£ĀņČ®ņŚÉ ļīĆĒĢ£ ņé┤ņäĀņČ® ĒÖ£ņä▒ ņŗżĒŚśņŚÉ ņé¼ņÜ®Ē¢łļŹś ņĪ░Ļ▒┤Ļ│╝ Ļ░Öņ£╝ļ®░, ņäĀņČ® ņĢī ļČĆĒÖöņ£©ņØĆ ņŗ£ļŻīņ▓śļ”¼ Ēøä 3ņØ╝, 7ņØ╝, 9ņØ╝ņ¦Ė ņĪ░ņé¼ĒĢśņśĆļŗż.
ņé┤ņäĀņČ®ņ£©(%)=[(ņ▓śļ”¼ĻĄ¼ņØś ņ╣śņé¼ņ£©ļīĆņĪ░ĻĄ¼ņØś- ņ╣śņé¼ņ£©)/(100ļīĆņĪ░ĻĄ¼ņØś-ņ╣śņé¼ņ£©)]├Ś100
ņĢī ļČĆĒÖöņ£©(%)=[ņ£ĀņČ®ņØś ņłś/(ņĢīņØś ņłś+ņ£ĀņČ®ņØś ņłś)]├Ś100
ļČäļ”¼ ĻĘĀņŻ╝ņØś ĒśĢĒā£ĒĢÖņĀü ļ░Å ļČäņ×ÉĻ│äĒåĄĒĢÖņĀü ļČäņäØ
ņäĀļ░£ļÉ£ KRA15-528 ĻĘĀņŻ╝ņØś ĒśĢĒā£ĒĢÖņĀü ĒŖ╣ņä▒ņØĆ BennettŌĆģŌĆÖs ĒĢ£ņ▓£ļ░░ņ¦ĆņŚÉ ņĀæņóģĒĢśņŚ¼ 28┬░C ļ░░ņ¢æĻĖ░ņŚÉņä£ 10ņØ╝ ļÅÖņĢł ļ░░ņ¢æĒĢ£ ļÆż Ļ┤Ćņ░░ĒĢśņśĆļŗż. ņŻ╝ņé¼ņĀäņ×ÉĒśäļ»ĖĻ▓ĮņØä ĒåĄĒĢśņŚ¼ ĻĘĀņŻ╝ņØś Ēżņ×ÉĒśĢĒā£ņÖĆ Ēü¼ĻĖ░ļź╝ Ļ┤Ćņ░░ĒĢśĻĖ░ ņ£äĒĢ┤ ļ®ĖĻĘĀļÉ£ ĻĖłņåŹņØä BennettŌĆģŌĆÖs ĒĢ£ņĀäļ░░ņ¦ĆņŚÉ ļ╣äņŖżļō¼Ē׳ ļü╝ņøī ļäŻņØĆ ļÆż ĻĖłņåŹĻ│╝ ļ░░ņ¦ĆĻ░Ć ļ¦×ļŗ┐ļŖö ņäĀņØä ļö░ļØ╝ KRA15-528 ĻĘĀņŻ╝ļź╝ ļÅäļ¦ÉĒĢśņśĆļŗż. 28┬░CņŚÉņä£ 10ņØ╝Ļ░ä ļ░░ņ¢æĒĢśņŚ¼ ĻĖłņåŹņØä ĒāĆĻ│Ā ņś¼ļØ╝ņś© ĻĘĀņ▓┤ļź╝ ņŻ╝ņé¼ņĀäņ×ÉĒśäļ»ĖĻ▓Į(MIRA3; TESCAN, Seoul, Korea)ņØä ĒåĄĒĢ┤ Ļ┤Ćņ░░ĒĢśņśĆļŗż(WilliamsņÖĆ Davies, 1967).
ļČäļ”¼ ĻĘĀņŻ╝ņØś 16S rRNA ņ£ĀņĀäņ×É ņŚ╝ĻĖ░ņä£ņŚ┤ņØä ļ░öĒāĢņ£╝ļĪ£ ļČäņ×ÉĻ│äĒåĄĒĢÖņĀü ļČäņäØņØä ņŗżņŗ£ĒĢśņśĆļŗż. KRA15-528ņØä BennettŌĆÖs ņĢĪņ▓┤ļ░░ņ¦Ć 5 mlņŚÉ ņĀæņóģĒĢśņŚ¼ 28┬░C, 150 rpm ņĪ░Ļ▒┤ņ£╝ļĪ£ 3ņØ╝Ļ░ä ļ░░ņ¢æĒĢśņśĆļŗż. ļ░░ņ¢æņĢĪņØä 4┬░C, 6,500g ņĪ░Ļ▒┤ņ£╝ļĪ£ 10ļČäĻ░ä ņøÉņŗ¼ļČäļ”¼ĒĢśņŚ¼ ņ¢╗ņØĆ ĻĘĀņ▓┤ļĪ£ļČĆĒä░ cetyltrimethylammonium bromideļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ genomic DNAļź╝ ņČöņČ£ĒĢśņśĆļŗż(TripathiņÖĆ Rawal, 1998). 16S rRNA ļČĆļČäņØś ņ”ØĒÅŁņØä ņ£äĒĢśņŚ¼ universal primer set (27F, 5ŌĆÖ-AGAGTTTGATCMTGGCTCAG-3ŌĆÖ; 1492R, 5ŌĆÖ-TACGGYTACCTTGTTACGACTT-3ŌĆÖ)ļź╝ ņØ┤ņÜ®ĒĢśņśĆņ£╝ļ®░, ņżæĒĢ® ĒÜ©ņåī ņŚ░ņćäļ░śņØæņØĆ Ex Taq polymerase (Takara, Ohtsu, Japan)ļź╝ ņé¼ņÜ®ĒĢśņŚ¼ 94┬░C 5ļČä 1ĒÜī, 94┬░C 30ņ┤ł, 58┬░C 30ņ┤ł, 72┬░C 30ņ┤łņŚÉņä£ 34ĒÜī, 72┬░CņŚÉņä£ 7ļČäĻ░ä 1ĒÜī ļ░śņØæņŗ£ņ╝░ļŗż. ĒÖĢņØĖļÉ£ ļ░┤ļō£ļŖö GeneAll ExpinŌäó PCR purification kit (GeneAll, Seoul, Korea)ņØś ļ░®ļ▓ĢņŚÉ ļö░ļØ╝ ņĀĢņĀ£ĒĢ£ Ēøä ņŚ╝ĻĖ░ņä£ņŚ┤ ļČäņäØņØä ņ£äĒĢ┤ ņŗ£ĒĆĆņŗ▒ ļČäņäØ ņä£ļ╣äņŖż(Macrogen, Daejeon, Korea)ļź╝ ņØ┤ņÜ®ĒĢśņśĆļŗż. ļČäņäØļÉ£ ņŚ╝ĻĖ░ņä£ņŚ┤ņØĆ BLASTn (http://www.ncbi.nlm.nih.gov) ĒöäļĪ£ĻĘĖļשņØä ĒåĄĒĢ┤ ļ╣äĻĄÉ ļČäņäØļÉśņŚłļŗż. ļČäņ×É Ļ│äĒåĄĒĢÖņĀü Ļ│äĒåĄņłśņØś ņ×æņä▒ņØĆ MEGA ĒöäļĪ£ĻĘĖļש Ēī©Ēéżņ¦Ćļź╝ ņé¼ņÜ®ĒĢśņŚ¼ ņłśĒ¢ēĒĢśņśĆņ£╝ļ®░(Tamura ļō▒, 2013), ļČäņäØņØä ņ£äĒĢ£ ņŚ╝ĻĖ░ņä£ņŚ┤ ņĀĢļĀ¼ņØĆ CLUSTAL W (Larkin ļō▒, 2007)ļź╝ ņØ┤ņÜ®ĒĢśņśĆļŗż. ļČäņ×ÉĻ│äĒåĄļÅäļŖö Ļ▒░ļ”¼Ē¢ēļĀ¼ļ▓Ģ ņżæņØś ĒĢśļéśņØĖ neighbor-joining ļ░®ļ▓Ģ(SaitouņÖĆ Nei, 1987)ņØä ņé¼ņÜ®ĒĢśņŚ¼ ĻĄ¼ņä▒ĒĢśņśĆĻ│Ā, bootstrap ļČäņäØņØĆ 1,000ĒÜī ņłśĒ¢ēĒĢśņśĆļŗż.
ļ░░ņ¢æņŚ¼ņĢĪ ņÜ®ļ¦ż ļČäĒÜŹļōżņØś ņé┤ņäĀņČ® ĒÖ£ņä▒
ļ░®ņäĀĻĘĀ ļ░░ņ¢æņŚ¼ņĢĪņØś ņÜ®ļ¦ż ļČäĒÜŹņŚÉ ļö░ļźĖ ņé┤ņäĀņČ® ĒÖ£ņä▒ņØä ņĪ░ņé¼ĒĢśĻĖ░ ņ£äĒĢ┤ ļ░░ņ¢æņŚ¼ņĢĪĻ│╝ ļÅÖļ¤ēņØś ņŚÉĒŗĖ ņĢäņäĖĒģīņØ┤ĒŖĖņÖĆ ļČĆĒāäņś¼ņØä ņØ┤ņÜ®ĒĢśņŚ¼ ņł£ņ░©ņĀüņ£╝ļĪ£ Ļ░üĻ░ü 2ĒÜīņö® ņÜ®ļ¦ż ļČäĒÜŹĒĢśņŚ¼ Ļ░ÉņĢĢ ļåŹņČĢĒĢśņśĆļŗż. ņŚÉĒŗĖ ņĢäņäĖĒģīņØ┤ĒŖĖ, ļČĆĒāäņś¼, ļ¼╝ļĪ£ļČĆĒä░ ņČöņČ£ļ¼╝ 0.01, 0.45, 0.59 gņö®ņØä ĒÖĢļ│┤ĒĢśņśĆņ£╝ļ®░, Ļ░ü ņČöņČ£ļ¼╝ņØä 100 mg/ml ļåŹļÅäĻ░Ć ļÉśļÅäļĪØ ļ®öĒāäņś¼(ņŚÉĒŗĖ ņĢäņäĖĒģīņØ┤ĒŖĖņÖĆ ļČĆĒāäņś¼ ņČöņČ£ļ¼╝) ļśÉļŖö 50% ļ®öĒāäņś¼(ļ¼╝ ņČöņČ£ļ¼╝)ņŚÉ ņÜ®ĒĢ┤ĒĢśņśĆļŗż. 96-well tissue culture plateņØś Ļ░ü wellņŚÉ ļ┐īļ”¼Ēś╣ņäĀņČ® 2ļĀ╣ ņ£ĀņČ®(ņĢĮ 50ļ¦łļ”¼/99 ╬╝l)ņØä ņ╣śņāüĒĢ£ Ēøä Ļ░üĻ░üņØś ņŗ£ļŻīļź╝ ņ▓śļ”¼ĒĢśņŚ¼ ņĄ£ņóģ ļåŹļÅäĻ░Ć 250, 500, 1,000 ┬Ąg/mlĻ░Ć ļÉśļÅäļĪØ ĒĢśņśĆļŗż. ļ¼┤ņ▓śļ”¼ ļīĆņĪ░ĻĄ¼ļĪ£ļŖö 1% ļ®öĒāäņś¼ņØä ņé¼ņÜ®ĒĢśņśĆņ£╝ļ®░, ņŗżĒŚśņØĆ 3ĒÜī ļ░śļ│Ą ņłśĒ¢ēļÉśņŚłļŗż. ņŗ£ļŻī ņ▓śļ”¼ Ēøä 48ņŗ£Ļ░ä ļÆż, ņé┤ņäĀņČ®ļźĀņØä ņĪ░ņé¼ĒĢśņśĆļŗż.
In vivo ļ┐īļ”¼Ēś╣ņäĀņČ®ļ│æ ļ░®ņĀ£ ĒÖ£ņä▒ Ļ▓ĆņĀĢ
ņśżņØ┤ļź╝ ĻĖ░ņŻ╝ļĪ£ ņØ┤ņÜ®ĒĢśņŚ¼ KRA15-528 ĻĘĀņŻ╝ ļ░░ņ¢æņŚ¼ņĢĪņØś ļ┐īļ”¼Ēś╣ņäĀņČ®ļ│æ ļ░®ņĀ£ĒÖ£ņä▒ņØä ņĪ░ņé¼ĒĢśņśĆļŗż. ņś©ņŗż(25┬░C-5┬░C)ņŚÉņä£ 5ņØ╝ ļÅÖņĢł ĒéżņÜ┤ ŌĆśļ░▒ļ»ĖļŗżļŗżĻĖ░ŌĆÖ ĒÆłņóģņØś ņśżņØ┤ļź╝ ļ®ĖĻĘĀĒĢ£ ļ¬©ļל 100 gņØ┤ ļŗ┤ĻĖ┤ ņ¦üĻ▓Į 7 cmņØś ĒöīļØ╝ņŖżĒŗ▒ ĒżĒŖĖņŚÉ ņØ┤ņŗØĒĢśņśĆļŗż. ĒåĀļ¦łĒåĀņŚÉņä£ ņ”ØņŗØļÉ£ ļ┐īļ”¼Ēś╣ņäĀņČ® ņĢīņØä ņłśĻ▒░ĒĢśņŚ¼ Ļ░ü ĒżĒŖĖļŗ╣ 5,000ņĢīņØä ņĀæņóģĒĢśņśĆļŗż. ņĀæņóģ Ēøä ĒĢ£ ņŗ£Ļ░ä ļÆż, KRA15-528 ĻĘĀņŻ╝ņØś ļ░░ņ¢æņŚ¼ņĢĪņØä ņśżņØ┤ ļ┐īļ”¼ ņŻ╝ļ│ĆņŚÉ 10 mlņö® Ļ┤ĆņŻ╝ņ▓śļ”¼ĒĢśņśĆņ£╝ļ®░, ņØ╝ņŻ╝ņØ╝ Ēøä Ļ░ÖņØĆ ļ░®ņŗØņ£╝ļĪ£ ļ░░ņ¢æņŚ¼ņĢĪņØä ĒĢ£ļ▓ł ļŹö ĒåĀņ¢æĻ┤ĆņŻ╝ ņ▓śļ”¼ĒĢśņśĆļŗż. ļīĆņĪ░ĻĄ¼ļĪ£ļŖö ņäĀņČ®Ēāä(a.i. 5% fosthiazate) 2,000ļ░░ ĒؼņäØņĢĪņØä 1ĒÜī ņ▓śļ”¼ĒĢśņśĆņ£╝ļ®░, ļ¼┤ņ▓śļ”¼ĻĄ¼ļĪ£ļŖö ļ®ĖĻĘĀņłśļź╝ ņØ┤ņÜ®ĒĢśņśĆļŗż. ņŗżĒŚśņØĆ Ļ░ü ņ▓śļ”¼ĻĄ¼ļŗ╣ 5Ļ░£ņØś ņŗØļ¼╝ņ▓┤ļź╝ ņé¼ņÜ®ĒĢśņŚ¼ 2ļ░śļ│ĄņØä ņłśĒ¢ēĒĢśņśĆĻ│Ā, ņäĀņČ® ņĢī ņĀæņóģ 2ņŻ╝ ĒøäņŚÉ ļ┐īļ”¼Ēś╣ ņ¦Ćņłśļź╝ ņĪ░ņé¼ĒĢśņśĆļŗż. ļ┐īļ”¼Ēś╣ ņ¦ĆņłśļŖö ņśżņØ┤ļ┐īļ”¼ņŚÉ ĒśĢņä▒ļÉ£ Ēś╣ņØś ņĀĢļÅäņŚÉ ļö░ļØ╝ 0-4ņØś ņ¦Ćņłśļź╝ ņé¼ņÜ®ĒĢśņŚ¼ 5ļŗ©Ļ│äļĪ£ ņĪ░ņé¼ĒĢśņśĆļŗż(0, 0%; 1, 1%-25%; 2, 26%-50%; 3, 51%-75%; 4, 76%-100%) (TaylorņÖĆ Sasser, 1978).
Ļ▓░Ļ│╝ ļ░Å Ļ│Āņ░░
ļ░®ņäĀĻĘĀņØś in vitro ņé┤ņäĀņČ® ĒÖ£ņä▒Ļ│╝ ņĢī ļČĆĒÖö ņ¢ĄņĀ£ ĒÖ£ņä▒
ņé┤ņäĀņČ® ĒÖ£ņä▒ņØä Ļ░Ćņ¦ĆļŖö ĻĘĀņŻ╝ņØś ņäĀļ░£ņØä ņ£äĒĢśņŚ¼ 670ņŚ¼ Ļ░£ņØś ļ░®ņäĀĻĘĀ ļ░░ņ¢æņŚ¼ņĢĪņØä ņØ┤ņÜ®ĒĢśņŚ¼ 96-well tissue culture plateņŚÉņä£ ņŖżĒü¼ļ”¼ļŗØņØä ņłśĒ¢ēĒĢśņśĆļŗż. ņĢĮ 670Ļ░£ņØś ļ░®ņäĀĻĘĀ ņżæ KRA15-528ņØ┤ Ļ░Ćņן ļåÆņØĆ ņé┤ņäĀņČ® ĒÖ£ņä▒ņØä Ļ░Ćņ¦ĆļŖö ĻĘĀņŻ╝ļĪ£ ņäĀļ░£ļÉśņŚłļŗż. KRA15-528 ĻĘĀņŻ╝ļź╝ ņäĀņČ® ĒśäĒāüņĢĪņŚÉ ņ▓śļ”¼ Ēøä, 48ņŗ£Ļ░äņ¦ĖņŚÉ Ļ┤Ćņ░░ĒĢśņśĆņØä ļĢī 10% ņ▓śļ”¼ĻĄ¼ņŚÉņä£ļŖö 71%, 5%ņŚÉņä£ļŖö 65%, 2.5%ņŚÉņä£ļŖö 61%ņØś ļåÆņØĆ ņé┤ņäĀņČ®ļźĀņØä ļ╣äļĪ»ĒĢśņŚ¼ ļåŹļÅä ņØśņĪ┤ņĀüņØĖ Ļ▓░Ļ│╝ļź╝ ļ│┤ņŚ¼ņŻ╝ņŚłļŗż. ļśÉĒĢ£ 1.25% ņ▓śļ”¼ĻĄ¼ņŚÉņä£ļÅä 19%ņØś ĒÖ£ņä▒ņØä ļéśĒāĆļāłļŗż(Fig. 1A). ļ░░ņ¢æņŚ¼ņĢĪ ņ▓śļ”¼ Ēøä, 72ņŗ£Ļ░ä ļ░Å 96ņŗ£Ļ░äņ¦ĖņŚÉņä£ļÅä 48ņŗ£Ļ░äņ¦Ė Ļ┤Ćņ░░ĒĢ£ ņé┤ņäĀņČ®ļźĀĻ│╝ ļ╣äņŖĘĒĢ£ ņé┤ņäĀņČ®ļźĀņØä ļéśĒāĆļé┤ņ¢┤ ņ¦ĆņåŹņĀüņØĖ ņé┤ņäĀņČ® ĒÖ£ņä▒ņØä ļ│┤ņ×äņØä ņĢī ņłś ņ׳ņŚłļŗż(data not shown). Jang ļō▒(2015)ņØĆ S. netropsis AN10065 ĻĘĀņŻ╝ņØś ļ░░ņ¢æņĢĪ ņČöņČ£ļ¼╝ņØä ņäĀņČ®ņŚÉ 5% ļåŹļÅäļĪ£ ņ▓śļ”¼ ņŗ£ 72ņŗ£Ļ░ä Ēøä 20% ņØ┤ĒĢśņØś ņé┤ņäĀņČ®ļźĀņØ┤ ļéśĒāĆļé¼ļŗżĻ│Ā ļ│┤Ļ│ĀĒĢśņśĆņ£╝ļ®░, Kim ļō▒(2011)ņØĆ S. sampsonii KK1024 ļ░░ņ¢æņĢĪņØä 10% ļåŹļÅäļĪ£ ņ▓śļ”¼Ē¢łņØä ļĢī 21%ņØś ņé┤ņäĀņČ®ļźĀņØä ļ│┤Ļ│ĀĒĢśĻ│Ā ņ׳ļŗż. ĒĢśņ¦Ćļ¦ī ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö KRA15-528 ĻĘĀņŻ╝ņØś ļ░░ņ¢æņŚ¼ņĢĪ 10% ņ▓śļ”¼ ņŗ£ 71%ņØś ļåÆņØĆ ņé┤ņäĀņČ® ĒÖ£ņä▒ņØä ĒÖĢņØĖĒĢśņśĆļŗż(Fig. 1A). ļ░░ņ¢æņĪ░Ļ▒┤ ļō▒ ņŗżĒŚśņĀü ĒÖśĻ▓ĮņØś ņ░©ņØ┤ļź╝ Ļ░ÉņĢłĒĢ£ļŗżĻ│Ā ĒĢśļŹöļØ╝ļÅä ņĢ×ņäĀ ņŚ░ĻĄ¼ Ļ▓░Ļ│╝ņÖĆ ļ╣äĻĄÉĒĢśņŚ¼ ļ│Ė ņŗżĒŚśņŚÉ ņé¼ņÜ®ļÉ£ Streptomyces sp. KRA15-528 ĻĘĀņŻ╝Ļ░Ć in vitroņŚÉņä£ ņÜ░ņłśĒĢ£ ņé┤ņäĀņČ®ļźĀņØä ļ│┤ņŚ¼ņŻ╝Ļ│Ā ņ׳ņØīņØä ņĢī ņłś ņ׳ņŚłļŗż.
Fig.┬Ā1
Effects of culture filtrates of Streptomyces flavogriseus KRA15-528 on J2 mortality (A) and egg hatching (B) of Meloidogyne incognita. Values are mean┬▒standard deviation of three replicates. Means with the same letter are not significantly different (P<0.05) according to DuncanŌĆÖs multiple range test. Abamectin (╬╝g/╬╝l) was used as a control. (A) The number of dead nematodes were counted 48 hours after treatment. Mortality rates (M) were corrected using AbbottŌĆÖs formula: M=[(Mt-Mc)/(100-Mc)]├Ś100, where Mt means mortality percentage in treatment and Mc means mortality percentage in control. (B) The number of hatched eggs were counted 3, 7, and 9 days after treatment. Egg hatch rate (EH) was calculated as follows: EH=[J/(E+J)]├Ś100, where J and E mean the juveniles and eggs of M. incognita, respectively.
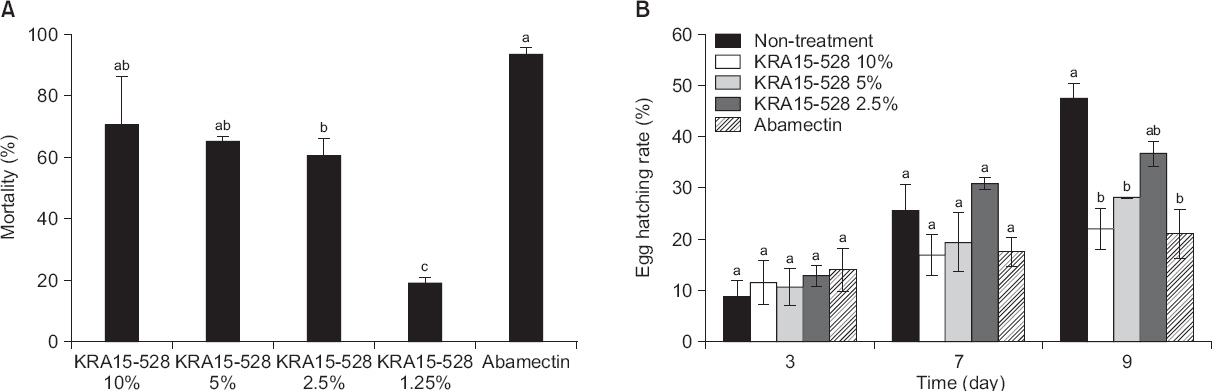
ņäĀņČ® ņĢī ļČĆĒÖöņ£©ņØĆ ņŗ£ļŻī ņ▓śļ”¼ Ēøä 7ņØ╝ņ¦ĖļČĆĒä░ ļ¼┤ņ▓śļ”¼ ļīĆņĪ░ĻĄ¼ņÖĆņØś ņ░©ņØ┤ļź╝ ļ│┤ņØ┤ĻĖ░ ņŗ£ņ×æĒĢśņśĆļŗż(Fig. 1B). ĒŖ╣Ē׳, ņŗ£ļŻīņ▓śļ”¼ Ēøä 9ņØ╝ņ¦Ė Ļ┤Ćņ░░ĒĢśņśĆņØä ļĢī, ļ¼┤ņ▓śļ”¼ĻĄ¼ņØś ņĢī ļČĆĒÖöņ£©ņØĆ 48%ņØĖ ļŹ░ ļ░śĒĢ┤, 10%ņÖĆ 5% ņ▓śļ”¼ĻĄ¼ņŚÉņä£ņØś ņĢī ļČĆĒÖöņ£©ņØĆ 22%ņÖĆ 28%ņśĆļŗż. ņ”ē, ļ░░ņ¢æņŚ¼ņĢĪ 10%ņÖĆ 5% ņ▓śļ”¼ĻĄ¼ļŖö ļ¼┤ņ▓śļ”¼ĻĄ¼ņÖĆ ļ╣äĻĄÉĒĢśņŚ¼ ņĢĮ 54%ņÖĆ 41%ņØś ņäĀņČ® ņĢī ļČĆĒÖö ņ¢ĄņĀ£ņ£©ņØä ļ│┤ņŚ¼ņŻ╝ņŚłļŗż(Fig. 1B). ņØ┤ņÖĆ Ļ░ÖņØĆ ņäĀņČ® ņĢī ļČĆĒÖö ņ¢ĄņĀ£ĒÜ©Ļ│╝ļŖö KRA15-528 ĻĘĀņŻ╝Ļ░Ć ņ╣£ĒÖśĻ▓Į ļ░®ņĀ£ņĀ£ļĪ£ņä£ Ļ░£ļ░£ Ļ░ĆļŖźĒĢ£ ņåīņ×¼ņ×äņØä ļéśĒāĆļé┤ņŻ╝ļŖö Ļ▓░Ļ│╝ļĪ£ ņāØĻ░üļÉśņŚłļŗż.
KRA15-528 ĻĘĀņŻ╝ņØś ĒśĢĒā£ĒĢÖņĀü ĒŖ╣ņä▒ ļ░Å Ļ│äĒåĄņłś ļČäņäØ
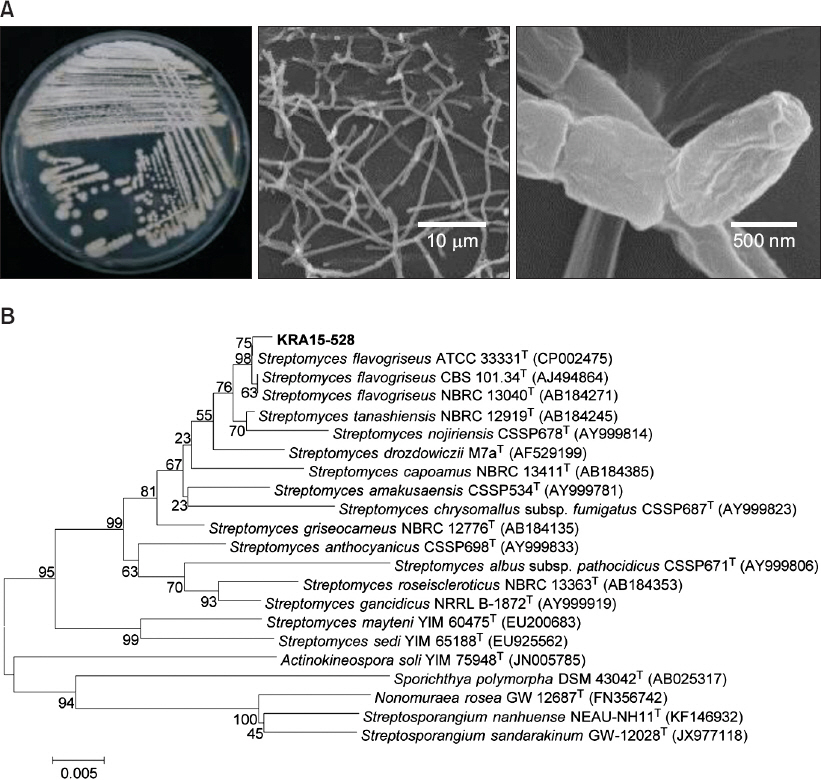
KRA15-528 ĻĘĀņŻ╝ļŖö BennettŌĆģŌĆÖs ĒĢ£ņ▓£ļ░░ņ¦ĆņŚÉ ļÅäļ¦ÉĒĢśņśĆņØä ļĢī ņé¼ņāüĒśĢņØś ĻĘĀņé¼ļŖö ĒÜīņāē ļ░Å ļģĖļ×ĆņāēņØä ļØĀļ®░, ņŻ╝ņé¼ņĀäņ×ÉĒśäļ»ĖĻ▓Įņ£╝ļĪ£ Ļ┤Ćņ░░ ņŗ£ ĻĘĀņé¼ņØś ļüØļČĆļČäņŚÉ ļČäņĀłļÉ£ ĒśĢĒā£ņØś ņøÉĒåĄ ļ¬©ņ¢æ Ēżņ×ÉĻ░Ć Ļ┤Ćņ░░ļÉśņŚłļŗż(Fig. 2A). Ēżņ×ÉņØś Ēü¼ĻĖ░ļŖö ņĢĮ 0.7-1 ╬╝mņśĆļŗż(Fig. 2A). 16S rRNA ņ£ĀņĀäņ×É ņŚ╝ĻĖ░ņä£ņŚ┤ņØä ņØ┤ņÜ®ĒĢ£ ļČäņ×É Ļ│äĒåĄĒĢÖņĀü ļČäņäØ Ļ▓░Ļ│╝, S. flavogriseus ATCC 33331Ļ│╝ 99.86%, S. tanashiensis NBRC 12919ņÖĆ 98.49%, S. nojiriensis CSSP678Ļ│╝ļŖö 96.83%ņØś ņŚ╝ĻĖ░ņä£ņŚ┤ ņ£Āņé¼ļÅäļź╝ ļ│┤ņśĆļŗż(Fig. 2B). ņØ┤ņŚÉ ņāüĻĖ░ņŚÉņä£ ļČäļ”¼ļÉ£ ĻĘĀņŻ╝ļź╝ S. flavogriseus KR15-528ļĪ£ ļ¬ģļ¬ģĒĢśņśĆļŗż.
Fig.┬Ā2
Morphological feature and phylogenetic analysis of Streptomyces flavogriseus KRA15-528. (A) Left picture shows a culture plate of KRA15-528 grown on BennettŌĆÖs agar medium 28┬░C for 10 days. Middle and right pictures are scanning electron micrograph images of hyphae and spore surface of KRA15-528, respectively. (B) Phylogenetic analysis was performed based on 16S rRNA gene sequence of KRA15-528 and 21 species of the genus Streptomyces. Neighbor-joining method was used for this analysis, and numbers at nodes indicate levels of bootstrap support (%) by 1,000 resampled datasets. National Center for Biotechnology Information accession numbers of each sequence are in parentheses. Bar, 5 substitution per 1,000 nt.
ņÜ®ļ¦ż ļČäĒÜŹļ│ä ņé┤ņäĀņČ® ĒÖ£ņä▒
KRA15-528 ĻĘĀņŻ╝ ļ░░ņ¢æņŚ¼ņĢĪņ£╝ļĪ£ļČĆĒä░ ņé┤ņäĀņČ® ĒÖ£ņä▒ ļ¼╝ņ¦łņØś ĒŖ╣ņä▒ņØä ĒīīņĢģĒĢśĻ│Āņ×É ņÜ®ļ¦ż ļČäĒÜŹļÉ£ ņŚÉĒŗĖ ņĢäņäĖĒģīņØ┤ĒŖĖ, ļČĆĒāäņś¼, ļ¼╝ņĖĄņØä Ļ░üĻ░ü ņäĀņČ® ĒśäĒāüņĢĪņŚÉ ņ▓śļ”¼ĒĢ┤ ļ│┤ņĢśļŗż. ņ▓śļ”¼ Ēøä 48ņŗ£Ļ░äņŚÉņä£ Ļ┤Ćņ░░ĒĢ£ Ļ▓░Ļ│╝, ņŚÉĒŗĖ ņĢäņäĖĒģīņØ┤ĒŖĖ ņČöņČ£ļ¼╝ ņ▓śļ”¼ĻĄ¼ņŚÉņä£ ļåÆņØĆ ņé┤ņäĀņČ® ĒÖ£ņä▒ņØ┤ ļéśĒāĆļé¼ļŗż(Fig. 3). ļČĆĒāäņś¼ ņČöņČ£ļ¼╝Ļ│╝ ļ¼╝ ņĖĄņØĆ 1,000 ┬Ąg/ml ļåŹļÅäļĪ£ ņ▓śļ”¼ ņŗ£ ņé┤ņäĀņČ®ļźĀņØ┤ Ļ░üĻ░ü 4%ņÖĆ 8%ļĪ£ ļ¦żņÜ░ ļé«ņĢśļŗż(Fig. 3). ņŚÉĒŗĖ ņĢäņäĖĒģīņØ┤ĒŖĖ ņČöņČ£ļ¼╝ņØś Ļ▓ĮņÜ░ 250, 500, 1,000 ┬Ąg/ml ņ▓śļ”¼ĻĄ¼ņŚÉņä£ Ļ░üĻ░ü 30%, 53%, 91%ņØś ļåŹļÅä ņØśņĪ┤ņĀüņØĖ ņé┤ņäĀņČ®ļźĀņØä ļ│┤ņŚ¼ņŻ╝ņŚłļŗż(Fig. 3). ļśÉĒĢ£, 72ņŗ£Ļ░ä ļ░Å 96ņŗ£Ļ░ä ĒøäņŚÉļÅä ņ¦ĆņåŹņĀüņØĖ ņé┤ņäĀņČ® ĒÜ©Ļ│╝ļź╝ ļ│┤ņŚ¼ņŻ╝ņŚłļŗż(data not shown). ņØ┤ļōż Ļ▓░Ļ│╝ļź╝ ļ░öĒāĢņ£╝ļĪ£, ņŚÉĒŗĖ ņĢäņäĖĒģīņØ┤ĒŖĖ ņČöņČ£ļ¼╝ņŚÉļŖö KRA15-528 ĻĘĀņŻ╝Ļ░Ć ņāØņä▒ĒĢśļŖö ņé┤ņäĀņČ® ĒÖ£ņä▒ ļ¼╝ņ¦łņØ┤ ĒżĒĢ©ļÉśņ¢┤ ņ׳ņ£╝ļ”¼ļØ╝ ņāØĻ░üļÉśļ®░, Ē¢źĒøä ņé┤ņäĀņČ® ĒÖ£ņä▒ ļ¼╝ņ¦łņØś ļÅÖņĀĢņŚÉ ļīĆĒĢ£ ņŚ░ĻĄ¼Ļ░Ć ĒĢäņÜöĒĢśļŗż.
Fig.┬Ā3
Nematicidal activity of two organic layers and aqueous layer from culture filtrate of Streptomyces flavogriseus KRA15-528. The number of dead nematodes were counted 48 hours after treatment. Mortality rate (M) was calculated as follows: M=[(Mt-Mc)/(100-Mc)]├Ś100, where Mt means mortality percentage in treatment and Mc means mortality percentage in control. Values are mean┬▒standard deviation of three replicates. Abamectin (╬╝g/╬╝l) was used as a control.
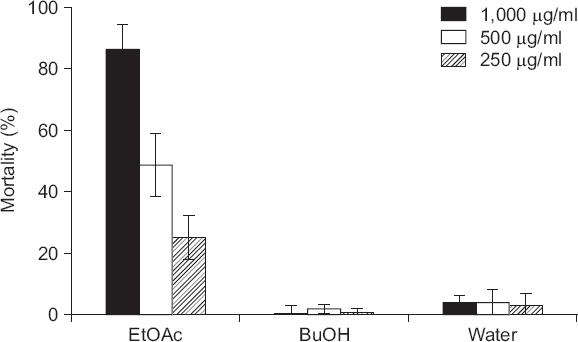
Ēśäņ×¼ ļīĆĒæ£ņĀüņØĖ ņé┤ņäĀņČ®ņĀ£ļĪ£ ĒīÉļ¦żļÉśĻ│Ā ņ׳ļŖö abamectinņØĆ avermectins B1aņÖĆ B1bņØś Ēś╝ĒĢ®ņĀ£ņØ┤ļ®░, ņØ┤ļōż ļ¼╝ņ¦łļōżņØĆ S. avermitilisņØś ņØ┤ņ░©ļīĆņé¼ņé░ļ¼╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż(LasotaņÖĆ Dybas, 1991; RobertsņÖĆ Hutson, 1999). ļśÉĒĢ£, KRA15-528 ĻĘĀņŻ╝ņÖĆ ļÅÖņØ╝ ņóģņØĖ S. flavogriseusļŖö protease, polycyclic xanthoneĻ│äņØś ĒĢŁņāØļ¼╝ņ¦łņØĖ SIIA-C2191AņÖĆ SIIA-C2191B ļō▒ņØś ņØ┤ņ░©ļīĆņé¼ņé░ļ¼╝ņØä ņāØņé░ĒĢ£ļŗżļŖö ļ│┤Ļ│ĀĻ░Ć ņ׳ņ£╝ļ®░, S. flavogriseus ACTK2 ĻĘĀņŻ╝ņØś ļČĆĒāäņś¼ ņČöņČ£ļ¼╝ņØĆ Staphylococcus aureus, Bacillus subtilis, Enterococcus aerogenes, Fusarium proliferatum ļō▒ņŚÉ ļīĆĒĢśņŚ¼ ĒĢŁĻĘĀĒÖ£ņä▒ņØä Ļ░Ćņ¦äļŗżļŖö ņŚ░ĻĄ¼Ļ▓░Ļ│╝Ļ░Ć ļ│┤Ļ│ĀļÉśņ¢┤ ņ׳ļŗż(DezfullyņÖĆ Ramanayaka, 2015; Ghorbel ļō▒, 2014; Zhao ļō▒, 2003). ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ņäĀļ░£ļÉ£ S. flavogriseus KRA15-528 ĻĘĀņŻ╝ļŖö ļ▓╝ ņ×Äņ¦æļ¼┤ļŖ¼ļ¦łļ”äļ│æ, ĒåĀļ¦łĒåĀ ņ×┐ļ╣øĻ│░ĒīĪņØ┤ļ│æ, ĒåĀļ¦łĒåĀ ņŚŁļ│æ, ļ░Ć ļČēņØĆļģ╣ļ│æ, Ļ│ĀņČö ĒāäņĀĆļ│æ ļō▒ņŚÉ ļ░®ņĀ£ĒÖ£ņä▒ņØ┤ ņŚåļŖö Ļ▓āņ£╝ļĪ£ Ļ┤Ćņ░░ļÉśņŚłņ¦Ćļ¦ī(data not shown), ĒĢŁĻĘĀĒÖ£ņä▒ņŚÉ ļīĆĒĢ£ ļ░®ņäĀĻĘĀņØś ļäōņØĆ ņŖżĒÄÖĒŖĖļ¤╝ņØä Ļ│ĀļĀżĒĢ£ļŗżļ®┤ ļŗżņ¢æĒĢ£ ņŗØļ¼╝ļ│æņøÉĻĘĀņØä ņØ┤ņÜ®ĒĢ£ ļ░®ņĀ£ĒÖ£ņä▒ņØś ņĪ░ņé¼Ļ░Ć ĒĢäņÜöĒĢśļ”¼ļØ╝ ņāØĻ░üļÉ£ļŗż.
ļ░®ņäĀĻĘĀņØĆ ļŗżņ¢æĒĢ£ ņāØļ”¼ĒÖ£ņä▒ļ¼╝ņ¦łņØä ņāØņé░ĒĢ£ļŗż. ņØ┤ļ¤¼ĒĢ£ ļ¼╝ņ¦łņØĆ Ēü¼Ļ▓ī ņäĖĒżļé┤ ļ¼╝ņ¦łĻ│╝ ņäĖĒżņÖĖ ļ¼╝ņ¦łļĪ£ ļéśļłī ņłś ņ׳ļŗż. Ēśäņ×¼Ļ╣īņ¦Ć ļ│┤Ļ│ĀļÉ£ ļ░®ņäĀĻĘĀņŚÉ ņØśĒĢ┤ ņāØņé░ļÉśļŖö ņé┤ņäĀņČ® ļ¼╝ņ¦łļōż, ņśłļź╝ ļōżļ®┤, avermectins, bafilomycin, lecanicidins ļō▒ņØĆ ņäĖĒżļé┤ ļ¼╝ņ¦łļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż(Burg ļō▒, 1979; Lacey ļō▒, 1995; Nonaka ļō▒, 2000). ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ S. flavogriseus KRA15-528 ĻĘĀņŻ╝ņØś ļ░░ņ¢æņŚ¼ņĢĪņ£╝ļĪ£ļČĆĒä░ ĒÖĢņØĖļÉ£ ņé┤ņäĀņČ® ĒÖ£ņä▒ņØĆ ĒÖ£ņä▒ļ¼╝ņ¦łņØ┤ ņäĖĒżņÖĖ ļ¼╝ņ¦łņØ╝ Ļ▓āņ£╝ļĪ£ ņČöņĀĢļÉśļ®░, ļÅÖņŗ£ņŚÉ ņāüĻĖ░ ĻĘĀņŻ╝ļĪ£ļČĆĒä░ ņé┤ņäĀņČ® ĒÖ£ņä▒ņØä Ļ░¢ļŖö ņäĖĒżļé┤ ļ¼╝ņ¦łņØä ļÅÖņĀĢĒĢ£ļŗżļ®┤ Ļ│ĀĒÖ£ņä▒ņØä ļ│┤ņØ┤ļŖö ļ»ĖņāØļ¼╝ ņé┤ņäĀņČ®ņĀ£ Ļ░£ļ░£ņØ┤ Ļ░ĆļŖźĒĢ┤ņ¦Ćļ”¼ļØ╝ ņāØĻ░üļÉ£ļŗż.
ĻĘĀņŻ╝ ļ░░ņ¢æņŚ¼ņĢĪņØś ļ┐īļ”¼Ēś╣ņäĀņČ®ņŚÉ ļīĆĒĢ£ in vivo ņŗżĒŚś
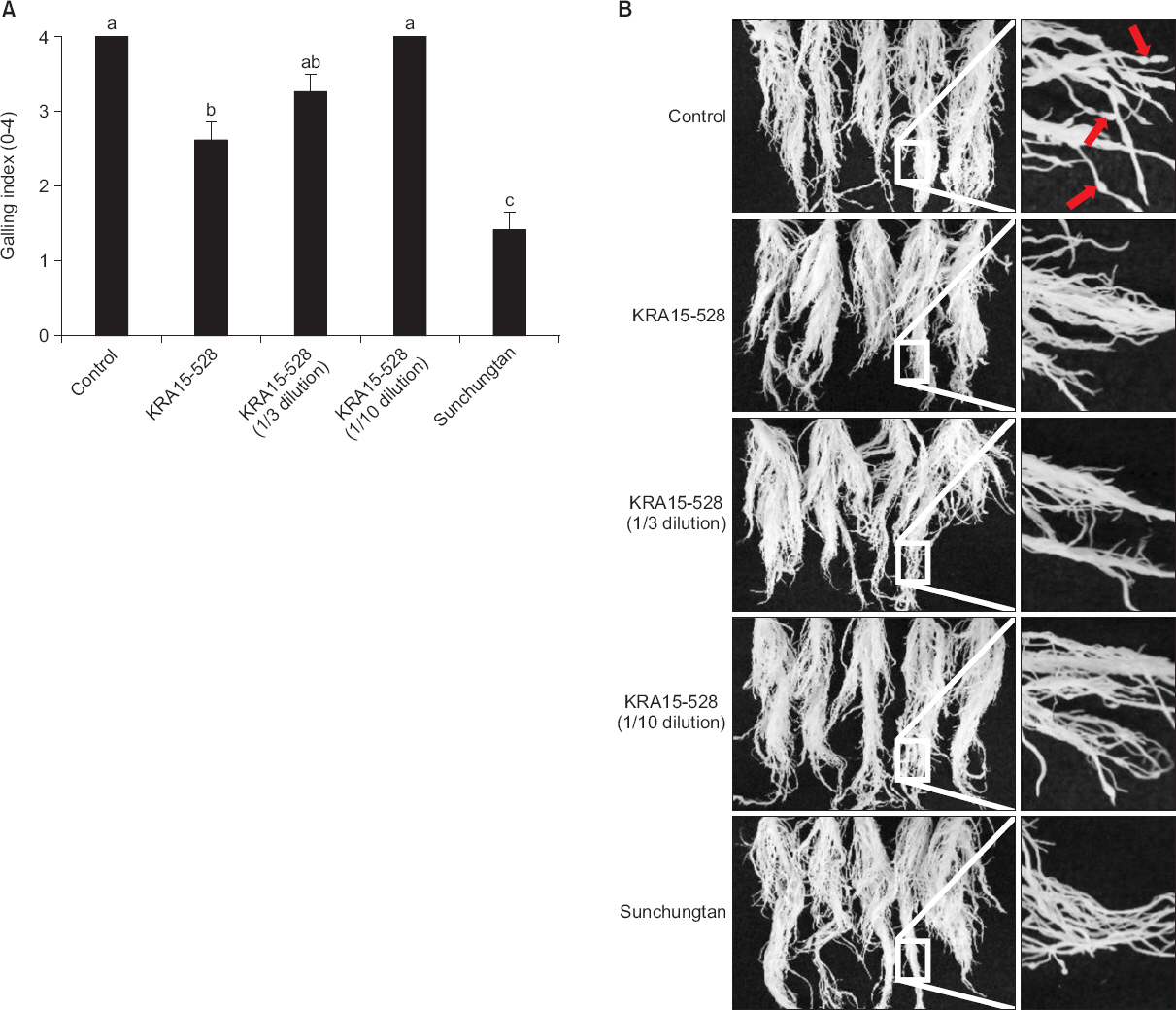
KRA15-528 ĻĘĀņŻ╝Ļ░Ć ļ┐īļ”¼Ēś╣ ņäĀņČ®ņŚÉ ļīĆĒĢ┤ ņŗØļ¼╝ļ│æļ░®ņĀ£ ĒÖ£ņä▒ņØä Ļ░Ćņ¦ĆļŖöņ¦Ć ņĢīņĢäļ│┤ĻĖ░ ņ£äĒĢśņŚ¼ ņśżņØ┤ņŚÉ ņäĀņČ®ņØä ņ▓śļ”¼ĒĢ£ Ēøä KRA15-528 ĻĘĀņŻ╝ņØś ļ░░ņ¢æņŚ¼ņĢĪņØä ņ▓śļ”¼ĒĢśņśĆļŗż. ļ░░ņ¢æņŚ¼ņĢĪņØś ņøÉņĢĪ, 1/3ļ░░, 1/10ļ░░ ĒؼņäØņĢĪ ņ▓śļ”¼ĻĄ¼ņŚÉņä£ Ļ░üĻ░ü 2.6, 3.3, 4.0ņØś ļ┐īļ”¼Ēś╣ĒśĢņä▒ņ¦Ćņłśļź╝ ļ│┤ņśĆņ£╝ļ®░, ļ¼┤ņ▓śļ”¼ĻĄ¼ņŚÉņä£ļŖö 4.0ņØś ļ┐īļ”¼Ēś╣ĒśĢņä▒ ņ¦Ćņłśļź╝ ļéśĒāĆļé┤ņŚłļŗż. KRA15-528 ļ░░ņ¢æņŚ¼ņĢĪ(ņøÉņĢĪ)ņØĆ ļ¼┤ņ▓śļ”¼ĻĄ¼ņÖĆ ņ£ĀņØśņĀüņ£╝ļĪ£ ņ░©ņØ┤ļź╝ ļ│┤ņśĆņ£╝ļéś, 1/3ļ░░ņÖĆ 1/10ļ░░ ĒؼņäØņĢĪņØĆ ņ£ĀņØśņĀüņØĖ ņ░©ņØ┤Ļ░Ć ņŚåņŚłļŗż(Fig. 4A) (TaylorņÖĆ Sasser, 1978). ļśÉĒĢ£, ņśżņØ┤ ņØ┤ņÖĖņŚÉļÅä ĒåĀļ¦łĒåĀļź╝ ļīĆņāüņ£╝ļĪ£ ĒĢ£ ņŗżĒŚśĻ▓░Ļ│╝ņŚÉņä£ļÅä KRA15-528 ĻĘĀņŻ╝ ļ░░ņ¢æņŚ¼ņĢĪņŚÉ ņØśĒĢ£ ļ░®ņĀ£ĒÖ£ņä▒ņØ┤ ĒÖĢņØĖļÉśņŚłņ£╝ļ®░, ļ¬©ļōĀ in vivo ņŗżĒŚśņāüņØś ņ▓śļ”¼ĻĄ¼ņŚÉņä£ ņĢĮĒĢ┤ļŖö ļ░£Ļ▓¼ļÉśņ¦Ć ņĢŖņĢśļŗż(data not shown).
Fig.┬Ā4
Inhibition activity of Streptomyces flavogriseus KRA15-528 to gall formation on cucumber roots. (A) At two weeks after infection of root-knot nematodes, the galling index was assessed according to a 0-4 galling scale; 0=0%, 1=1%-25%, 2=26%-50%, 3=51%-75%, and 4=76%-100% (Taylor and Sasser, 1978). Each value represents the mean┬▒standard deviation of five replications. The experiment was performed twice with similar results (data from a representative experiment are shown). Means with the same letter are not significantly different (P<0.05) according to DuncanŌĆÖs multiple range test. (B) Galling formation was suppressed by treatment of culture filtrate of KRA15-528. Photographs were taken 2 weeks after inoculation of Meloidogyne incognita. Left panel is magnified pictures of right panel, and red arrows indicate galls formed at roots.
ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö 670ņŚ¼ Ļ░£ņØś ļ░®ņäĀĻĘĀ Ļ░ĆņÜ┤ļŹ░ Ļ░Ćņן ņé┤ņäĀņČ® ĒÖ£ņä▒ņØ┤ ņÜ░ņłśĒĢ£ S. flavogriseus KRA15-528 ĻĘĀņŻ╝ļź╝ ņäĀļ░£ĒĢśņśĆĻ│Ā, ļ│Ė ĻĘĀņŻ╝ņØś ņé┤ņäĀņČ® ĒÖ£ņä▒ ļ¼╝ņ¦łņØĆ ņŚÉĒŗĖ ņĢäņäĖĒģīņØ┤ĒŖĖļĪ£ ņČöņČ£ņØ┤ Ļ░ĆļŖźĒĢ£ ņäĖĒżņÖĖ ļ¼╝ņ¦łļĪ£ ĒÖĢņØĖļÉśņŚłļŗż. ļśÉĒĢ£ ņśżņØ┤ļź╝ ņØ┤ņÜ®ĒĢ£ ņś©ņŗż ņŗ£ĒŚśņŚÉņä£ļÅä ļ┐īļ”¼Ēś╣ņäĀņČ®ņŚÉ ļīĆĒĢśņŚ¼ ļ░®ņĀ£ ĒÖ£ņä▒ņØä ļ│┤ņśĆļŗż. Ē¢źĒøä, ļ│Ė ĻĘĀņŻ╝ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ļ»ĖņāØļ¼╝ ņé┤ņäĀņČ®ņĀ£ļź╝ Ļ░£ļ░£ĒĢśĻĖ░ ņ£äĒĢ┤ņä£ļŖö ņé┤ņäĀņČ® ĒÖ£ņä▒ ļ¼╝ņ¦ł ļČäļ”¼ ļ░Å ĻĄ¼ņĪ░ ļÅÖņĀĢ, ņĄ£ņĀü ļ░£ĒÜ© Ļ│ĄņĀĢ, ņĄ£ņĀüņØś ņé┤ņäĀņČ® ĒÖ£ņä▒ņØä ņ£äĒĢ£ ņĀ£ņĀ£ Ļ░£ļ░£ ļō▒ņØś ņČöĻ░ĆņĀüņØĖ ņŚ░ĻĄ¼Ļ░Ć ĒĢäņÜöĒĢśļŗż.
ņÜöņĢĮ
ļ┐īļ”¼Ēś╣ņäĀņČ®ņŚÉ ņØśĒĢ£ ļ│æ ļ░£ņāØņØĆ ņ×æļ¼╝ņłśĒÖĢļ¤ēņØś Ļ░Éņåīļź╝ ņ┤łļלĒĢśņŚ¼ ņżæņÜöĒĢ£ ļ¼ĖņĀ£ļĪ£ ņØĖņŗØļÉśĻ│Ā ņ׳ļŗż. ĒÖöĒĢÖļåŹņĢĮņØś ņé¼ņÜ®ņØĆ ņäĀņČ®ļ░®ņĀ£ļź╝ ņ£äĒĢ┤ ĒÜ©Ļ│╝ņĀüņØĖ ļ░®ļ▓Ģ ņżæņØś ĒĢśļéśļĪ£ Ļ│ĀļĀżļÉśņ¦Ćļ¦ī ĒÖśĻ▓ĮņśżņŚ╝ņØ┤ļéś ņØĖņČĢņŚÉ ļīĆĒĢ£ ļÅģņä▒Ļ│╝ Ļ░ÖņØĆ ļ¼ĖņĀ£ļź╝ ņĢ╝ĻĖ░ĒĢśĻ│Ā ņ׳ļŗż. ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö ņ╣£ĒÖśĻ▓Į ņé┤ņäĀņČ®ņĀ£ Ļ░£ļ░£ņŚÉ ņé¼ņÜ®ļÉĀ ņłś ņ׳ļŖö ļ»ĖņāØļ¼╝ņØä ņäĀļ░£ĒĢśĻ│Āņ×É, ĒåĀņ¢æņ£╝ļĪ£ļČĆĒä░ ļČäļ”¼ĒĢ£ 670ņŚ¼ Ļ░£ņØś ļ░®ņäĀĻĘĀņØä ļīĆņāüņ£╝ļĪ£ ļ┐īļ”¼Ēś╣ņäĀņČ®ņŚÉ ļīĆĒĢ£ ņé┤ņäĀņČ® ĒÖ£ņä▒ņØä ņĪ░ņé¼ĒĢśņśĆļŗż. ņØ┤ļōż ņżæ Ļ░Ćņן ņÜ░ņłśĒĢ£ ņé┤ņäĀņČ® ĒÖ£ņä▒ņØä ļéśĒāĆļé┤ļŖö KRA15-528 ĻĘĀņŻ╝ļź╝ ņäĀļ░£ĒĢśņśĆņ£╝ļ®░, 16S rRNA ņ£ĀņĀäņ×É ņŚ╝ĻĖ░ņä£ņŚ┤ņŚÉ ĻĖ░ņ┤łĒĢśņŚ¼ S. flavogriseusļĪ£ ļÅÖņĀĢĒĢśņśĆļŗż. S. flavogriseus KRA15-28 ĻĘĀņŻ╝ ļ░░ņ¢æņŚ¼ņĢĪņØä ņĄ£ņóģļåŹļÅä 10%Ļ░Ć ļÉśļÅäļĪØ J2 ļŗ©Ļ│äņØś ņ£ĀņČ®Ļ│╝ ņäĀņČ® ņĢīņŚÉ Ļ░üĻ░ü ņ▓śļ”¼ ņŗ£, 48ņŗ£Ļ░ä Ēøä 71%ņØś ņäĀņČ® ņ╣śņé¼ņ£©Ļ│╝ 9ņØ╝ņ¦ĖņŚÉ 54%ņØś ņäĀņČ® ņĢī ļČĆĒÖö ņ¢ĄņĀ£ņ£©ņØä ĒÖĢņØĖĒĢśņśĆļŗż. ļ░░ņ¢æņŚ¼ņĢĪņØä ņŚÉĒŗĖ ņĢäņäĖĒģīņØ┤ĒŖĖ, ļČĆĒāäņś¼ņØä ņØ┤ņÜ®ĒĢśņŚ¼ ņÜ®ļ¦ż ļČäĒÜŹĒĢśņŚ¼ ņŚÉĒŗĖ ņĢäņäĖĒģīņØ┤ĒŖĖņÖĆ ļČĆĒāäņś¼, ļ¼╝ ņČöņČ£ļ¼╝ņŚÉ ļīĆĒĢ£ ņé┤ņäĀņČ® ĒÖ£ņä▒ņØä ņĪ░ņé¼ĒĢ£ Ļ▓░Ļ│╝, ņŚÉĒŗĖ ņĢäņäĖĒģīņØ┤ĒŖĖ ņČöņČ£ļ¼╝ļĪ£ļČĆĒä░ļ¦ī ļåÆņØĆ ņé┤ņäĀņČ® ĒÖ£ņä▒ņØä ĒÖĢņØĖĒĢśņśĆļŗż. 1,000, 500, 250 ┬Ąg/mlņØś ļåŹļÅäņŚÉņä£ Ļ░üĻ░ü 91%, 53%, 30%ņØś ņ╣śņé¼ņ£©ņØä ļéśĒāĆļé┤ņŚłļŗż. ņØ┤ņÖĖņŚÉļÅä, ņśżņØ┤ņŚÉ ļīĆĒĢ£ ļ┐īļ”¼Ēś╣ņäĀņČ®ļ│æ ļ░®ņĀ£ ĒÖ£ņä▒ņØä ņĪ░ņé¼ĒĢ£ Ļ▓░Ļ│╝, ļ░░ņ¢æņŚ¼ņĢĪ ņ▓śļ”¼ĻĄ¼ņŚÉņä£ ļłłņŚÉ ļØäĻ▓ī ļ┐īļ”¼Ēś╣ĒśĢņä▒ņØ┤ ņ¢ĄņĀ£ļÉśļŖö ĒśäņāüņØä Ļ┤Ćņ░░ĒĢśņśĆļŗż. ļö░ļØ╝ņä£, S. flavogriseus KRA15-528ņØĆ ļ┐īļ”¼Ēś╣ņäĀņČ®ļ│æņØä ļ░®ņĀ£ņŚÉ ņé¼ņÜ®ĒĢĀ ņłś ņ׳ļŖö ļ»ĖņāØļ¼╝ņĀ£ņĀ£ļĪ£ņä£ņØś Ļ░ĆļŖźņä▒ņØ┤ ņ׳ņØīņØä ĒÖĢņØĖĒĢĀ ņłś ņ׳ņŚłļŗż.
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






